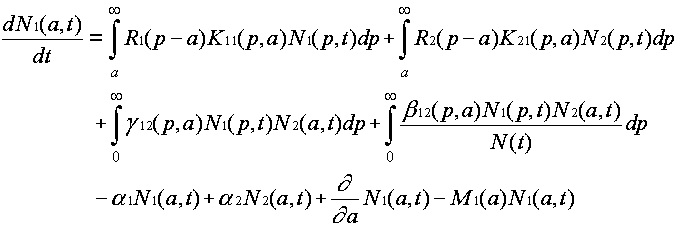 (1)
(1)
by Aaron Lynch
Note: This paper is an elaborated and refined successor to the published
peer-reviewed papers Lynch [1998] and Lynch [1991], and will itself be
sent for peer-reviewed publication. As a matter of record, the 2001 version
of this paper (as cited in other works) has been preserved at http://www.thoughtcontagion.com/UED2001.htm.
Likewise, a June, 2003 version of this paper has been preserved at
http://www.thoughtcontagion.com/UEDJune2003.htm.
The mathematical part of this paper may not display properly on all
versions of all web browsers and operating systems. The mathematical section
will only display properly if the following line contains the lower case
Greek letters Alpha, Beta, and Gamma, respectively:
a b g
Readers who do not wish to read differential equations should skip parts of section 12, "Population Dynamics."
An evolutionary replicator theory of mental/brain information is presented. With all replicator theories resting at least tacitly upon the fundamental notions of causation and of calling two or more entities "the same" with respect to an abstraction, the concept is rendered explicit in defining the terms memory items and thought contagions. It is argued that evolutionary cultural replicator theory may have no "absolute" system of memory abstractions much as physics has no absolute coordinate system (framework of space-time abstractions). A system of representing idea transistion events is introduced. The term "thought contagion" is given a technical definition. Arguments that sets of thought contagions are generally only partially ordered then provide a formal reason for not formally including "size" as a crucial element in defining units of replication. Differential equations are developed for thought contagion host population versus time in a two-variant system, modeling the dynamics whereby events at the individual level give rise to trends at the population level. This lays a foundation for computerized simulations and the falsification or validation of specific thought contagion hypotheses, and for testing quantitative evolutionary cultural replicator theory with animal experiments. As cultural replicator hypotheses generally involve observable communication events, they are found to have stronger empirical standing than hypotheses involving unidentified genes. Mechanisms of creativity as a population phenomenon are examined, with cultural evolutionary replicator analysis yielding a novel explanation for the temporal clustering of independent co-creations. Creation and propagation are integrated into a theory of evolution by variation and natural selection of cultural replicators.
KEYWORDS: Cultural replicator, evolution, replication, abstraction, transmissivity, receptivity, longevity, differential equation.
In his 1973 paper "Elementary Self-Replicating Instructions and their Works: Toward a Radical Reconstruction of General Anthropology Through a General Theory of Natural Selection," [Cloak, 1973] proposed and elaborated an evolutionary cultural replicator theory using symbolic analysis of replication event diagrams. His non-metaphoric analysis used a very broad definition of the term "instruction" that encompassed both neural features and features of genetic material. The formal proposal of this theory followed cultural microevolution studies conducted in a village of Trinidad during 1963 to 1965 that were the basis of Cloak's 1966 Ph.D dissertation [Cloak 1966a]. Early discussion of proposed "units of cultural instruction" and their self-propagating effects also appears in Cloak's short 1966 paper "Cultural Microevolution," [Cloak, 1966b].
Later, Cloak's more widely cited paper "Is a Cultural Ecology Possible" [Cloak, 1975] called attention to the 1973 paper, but did not fully inform readers of how extensively the theoretical framework and empirical work had been elaborated from 1966 to 1975. The wide awareness of a paper that, in its title, appeared merely to ask whether a cultural ecology was possible may have contributed to a wide presumption that technical elaboration of such a theory, did not actually exist or did not have any empirical basis. Subsequent popular works (e.g., Dawkins, 1976) that cited the 1975 paper without mentioning the 1973 paper may have contributed to the propagating misimpression that non-metaphoric technical work had not been done in this field [note 1]. For example, Cloak [1973] introduces the concept of a replicating unit, not using the word "unit" specifically, but rather, by referring to a replicator's unitary status with the word "elementary." He also formally states what makes a replicator (i.e., "self-replicating instruction") "elementary," which is to say, a unit. However, Dawkins [1976] and many subsequent works treat the "units," and by implication, the "elementary" aspects, as if it did not need careful definition or as if one could just assume that they were key features of culture. Section 8 of this paper discusses proposed unitary or elementary aspect of replicators in further detail.
Dawkins did co-edit a volume containing a subsequent non-metaphoric technical paper by Cloak [Cloak, 1986], but by that time, a great percentage of people aware of evolutionary cultural replicator theory had been convinced that the field arose by analogy and metaphor to genetics [note 2]. Making matters worse was the remarkable fact that Cloak's 1986 paper did not cite or mention his own 1973 paper, but described the 1986 work--instruction theory, event diagrams, and all--as being a formalization of later works by Dawkins. While the 1986 paper did contain new material, and did make clear that there are more kinds of instructions than the neural and genetic kinds discussed in the 1973 paper, it was not chronologically possible for the considerable body of instruction theory presented in both papers to have been simply a formalization of The Selfish Gene [Dawkins, 1976] and "Universal Darwinism" [Dawkins, 1983]. However, Cloak's own published statements of 1986 could subsequently be read as attributing the origins of evolutionary cultural replicator theory to Dawkins, even though this clearly was not the case. Meanwhile, a short discussion of the history of evolutionary cultural replicator theory in The Blind Watchmaker, [Dawkins, 1986, p. 157-158], can easily be read as misattributing the origins of evolutionary cultural replicator theory to The Selfish Gene [Dawkins, 1976]. While Cloak's 1986 chapter contains some speculations about early selection in macromolecules that could have benefited from collaboration with a physical or biochemical scientist, the overall paper was much more technically developed than Dawkins's works. By 1989, Dawkins had published a revised edition of The Selfish Gene [Dawkins, 1989], three years after he co-edited the volume containing Cloak [1986]. Yet the 1989 edition of The Selfish Gene made no mention of Cloak's 1986 paper, despite containing an updated bibliography and 11 pages of new endnotes to what was originally a 13-page chapter on cultural replicators in Dawkins [1976]. Thus, at least 12 years of academic and popular writing came to widely misconstrue evolutionary cultural replicator theory as originating in metaphor, popularization, and even word coinage, while most scientists remained unaware of the technical foundations of evolutionary cultural replicator theory.
Even if more people had been aware of Cloak's work, his term "instruction" may have had some limitations in generality as well as perhaps in self-explanation for interdisciplinary audiences. Cloak [1973] defines neural instructions in terms of neural structures that are replicated between organisms. A more general concept is that of a memory item that can be replicated between organisms. A memory item, defined with sufficient abstraction, need not require that "the same" piece of information be stored in "the same way" in different brains. Similarly, one can define units of computer code such as "bytes," "programs," etc. that need not be stored in the same way across different machines in order to be considered "the same." Most people already use abstract representations of memory content on a daily basis to discuss ideas. When we say that two people have "the same" idea, we do not use "sameness" to mean equality in every concrete detail, or even that we know that some structure is the same in different people. Otherwise, we would have trouble correctly saying that two people have "the same" idea. Perceiving two people to have "the same" idea involves abstracting out a set of common qualities. So saying that two people's ideas are "the same" only means that they are in some way "of the same kind."
Just because one can write out some statement does not mean that the statement represents a useful abstraction about real people in the population. Some statements are much more useful than others as representations of people's ideas. One can make up vastly more statements that do not usefully represent people's ideas than one can make up statements that do usefully represent people's ideas, even as there are an enormous number of statements that do usefully represent people's ideas. A nonsense statement such as "The air cracked off of the ruby" simply does not correspond usefully to an idea spreading in the population -- especially any idea spreading before the statement appeared in this paper. Some statements may correspond somewhat usefully to ideas spreading in real people, while other statements may correspond very usefully to ideas spreading in real people. This paper does not attempt to sort out the statements that correspond usefully to people's ideas from the statements that have no useful correspondence to people's ideas. Anyone who uses evolutionary epidemiology of ideas must therefore already have either a formal or an informal way of discerning statements that do correspond usefully to people's ideas from those that do not. For scientists, this may depend heavily upon what other theoretical frameworks they are using.
This ability to say that two people have the same idea lies at the foundation of the notion of a "replicating idea." When an idea "replicates," it acts to produce or preserve ideas that we call "the same idea." The resultant ideas can for now be called "self-replicated" ideas. To be a self-replicated idea means not only to have resulted from a given idea, but also to be "the same" as that idea. This is also spelled out symbolically in Cloak (1973). Specifically, it means meeting some abstract, observer-defined criterion for sameness.
Self-replicated ideas are not all exact replicas of their originals. A wide range of ideas may result from each self-propagating original. An observer just selectively lumps these proliferated ideas along with their original(s) into a set, using an abstract inclusion criterion.
As an example, the belief that "abortion is wrong" has a wide range of different meanings to different people. The range includes people who regard abortion as merely unethical to those who see the "morning after pill" as a high felony. So, although the belief varies greatly from person to person, its occurrences are all "the same" in the limited respect of fitting the above definition. Encountering a range of such beliefs in actual people, one "abstracts out" a common element running through all of them. On writing out a definition of this abstracted common element, or sameness criterion, one can proceed to use it for natural selection reasoning.
The abstraction could just as well be more general or more specific, depending upon our interests. One might, for instance, choose the more specific belief "abortion is a mortal sin." Then many quantitative variables involved in its natural selection would differ from those associated with the broader definition. First, the more restrictive definition would almost certainly identify a smaller host population. Second, when the hosts communicate their belief to friends and family, a likely smaller fraction of listeners will become new hosts per exposure. This is because any listeners who go away newly convinced that abortion is merely unethical no longer count as new hosts. Third, once a host, one may do less "ideological wavering" before being counted as a drop out. That is, remaining the host of a more restrictively defined belief takes more restrictive preservation. So changing an idea's definition can make a big difference in the evolutionary phenomena identified with it.
Distinguishing Neurally Stored Memory Items From Other Cultural Replicators
Many psychological and cultural phenomena other than ideas can be observed or considered to self-replicate. These include certain habits, attitudes, class identities, motor behaviors, verbal behaviors, cognitive associations, education, emotional dispositions, addictions, possession of artifacts, computer viruses, text contagions such as chain letters, and even neurotic and psychotic symptoms. Those who hold these acquired traits etc. need not be conscious of them in order for the acquired traits or other items to be analyzed for their self-replicating effects. For example, parents who have learned never to associate with "black people" may imprint this disposition upon their children by way of example display and imitation. The children, then, may continue not associating with "black people," and this social segregation can prevent them from learning that there is no good reason not to associate with "black people." One can thus define a non-association with "black people" trait that preserves itself by preventing the acquisition of information that would otherwise cause hosts to drop out of the non-associating trait. That, in turn, can preserve the idea long enough for children so imprinted to grow up and raise children of their own, and imprint the non-association trait on to a new generation. The word "idea" is used in a very general sense here, and does not indicate a conscious awareness nor explicit verbal communication. In the case of shunning people based on skin color, the learned trait, (called an "idea," in the general sense), may also give rise to rationalizations after the fact. Such rationalizations may then arise as consciously held consequences of unconscious early imprinting. Early imprinting may also render people more receptive or susceptible to rationalizations of racism and other unconsciously acquired traits. This can help the rationalizations spread as thought contagions in their own right, in addition to the numerous other mechanisms of racism and fascism contagion discussed in Lynch (2001b).
The wide range of traits covered in the branch of evolutionary cultural replicator analysis that constitutes the evolutionary epidemiology of ideas may be broadly classified as human memory content. This category is more general than the word "idea" or even "memory" usually connote. It includes everything in the fairly broad meaning of "memory," as succinctly defined in Merriam Webster's Collegiate Dictionary, 10th edition: "the store of things learned and retained from an organism's activity or experience as evidenced by modification of structure or behavior or by recall and recognition." Thus, the principal abstractions manipulated within the evolutionary epidemiology of ideas are memory abstractions. As used in this paper, "memory" does not include inanimate propagating items such as chain letters, Bibles, etc unless the context indicates otherwise. Nor does it include traits considered genetically instinctual unless otherwise indicated by the text. While many of these other propagating items may be considered to be or contain non-neurally stored memory, the term memory item in this paper is generally used to identify neurally-stored memory items. For brevity, the word "idea" is often used in this paper in a very general sense to indicate a neurally-stored memory item.
With neurally-stored memory items, the measurement of propagation is readily standardized in terms of a host count. Thus, a chain letter or the copying machine duplicating it do not count as hosts, but the person photocopying the letter can, if the letter was read, count as a host. The relationship of artifacts to ideas is discussed later. It should be noted, however, that the evolutionary epidemiology of ideas is not intended as a complete theory of cultural evolution. Chain letters, artifacts, behaviors, computer viruses, and so forth can be considered as replicators in their own right. For example, the directions in a chain letter might only exist one at a time in a person's short-term memory in order for the person to execute the full list of directions and thus pass on the letter. The full set of directions therefore might not occur in the person's nervous system even as the letter is copied and sent on to others. There might, however, be some co-propagating idea such as the promise of good luck for the sender that is passed in its complete form from one sender to another.
If an memory item resides very redundantly in someone's brain, that person still counts as only one host and one memory item instantiation. The number of duplicates of a memory item in one brain is not currently measurable, so it escapes further treatment in this article.
Representing Ideas Symbolically
Ideas can be represented conveniently with symbols such as "A," "B," etc. Thus, we can call the hell belief idea A, the imminent doomsday belief memory item B, and a combination of ideas such as the hell/imminent doomsday combination "A*B." The "*" indicates that A and B are instantiated in the same host. Extending this concept, one can represent a whole system of ideas as "A*B*C*..."
Symbolic expressions can also represent idea replication much the way
chemists represent chemical reactions. Thus, the hell belief's non-parental
conversion propagation may be represented as
A transition such as
It should be noted that events such as A
Even multistage evolutionary events are handily represented by this
system. For instance, the childbirth event can be extended to the form
Some multistage events are best represented by two or more diagrams.
For example, if two hosts of A have a baby non-host and then have a private
teacher instill the A-idea in the child, one would represent it as two
events:
Multi-idea events describe phenomenalike idea alteration or recombinationtoo
complex to represent with one complementary idea pair alone. For example,
forming the hell/imminent doomsday pair by recombination can be represented
as
Ideas A, B, and C are all rather "unpleasant" ideas, so it does not seem too surprising to find them propagating cooperatively. Yet in the real world, these three propagate cooperatively with a very "pleasant" idea D: "Love your neighbor as yourself." A*B*C*~D motivates its hosts to spread their faith only to those "unbelievers" that they really care to see saved from hell. In marked contrast, A*B*C*D hosts should be motivated to spread their faith to any unbelievers they should happen to meet. So the combination of ideas probably spreads more vigorously due to the inclusion of the D-idea.
When ideas propagate synergistically as do A, B, C, and D, they may propagate primarily as a set rather than individually. In such cases, the set may be usefully defined as one idea. Thus, one might define E = A*B*C*D and consider E to be a stable, propagating idea in its own right. This approach can sometimes be used to study very large ideological systems, such as religious and political doctrines, by treating them as single (but lengthily defined) memory abstractions. The propagation events, however, often contain many intricate stages.
Moreover, in specifying idea "X" by an equation like
Competition rather than cooperation characterizes many interacting ideas. An idea competes against its complement in the knowledge-of-birth-control case mentioned above. The two complementary ideaseach "armed" with distinct replication advantages"struggle" for host population.
Yet more heated than this competitionat least in some nationsis the competition between moral stances on the subject. Personal morality, after all, has a greater bearing on an adult's use or non-use of various methods in a society where birth control information is highly available. Moreover, morality strongly influences the parental decision to actively instruct children on birth control or to actively obstruct their learning on the subject.
So in addition to the knowledge-of-birth-control idea (idea A), we should
also consider idea B, the moral acceptance of practicing birth control,
and idea C, the belief that birth control is immoral. Here people are born
with the ~A*~B*~C combination, but they never end up simultaneously having
B and C later on. This is because B and C are contrary, as distinct
from complementary ideas. Since they are mutually exclusive, they are destined
to have a competitive rather than a cooperative relationship in the population.
The B idea is favored by a high occurrence of the non-parental conversion
event
Homogenic and Heterogenic Events
The birth control example as treated so far fails to acknowledge people's ability to independently invent or re-invent moral decisions on their own. People are portrayed as acquiring ideas only by copying them from others or by being born with them (as in the case of negatively defined memory items).
To remedy this omission, consider the cases of two students who learn
about birth control methods and then make moral judgments based on what
they have learned. The first decides the practice is morally acceptable
So one idea precipitates the formation of another idea, instead of just
a new copy of itself. Ideas that do this are called heterogenic,
or "other-forming." The corresponding events, such as
Idea transition event diagrams and terminology now acknowledge that
people can form opinions without simply copying others' ideas. Yet on many
topics, people copy more ideas than they either originate or "re-originate."
For many beliefs, such as the birth control taboo, homogenic formation
far outweighs heterogenic formation in its overall occurrence rate. The
reason is that homogenic events have the tendency toward self iteration
while heterogenic events do not. Generally speaking, anything that increases
the availability of the input ideas will increase the expected frequency
of the event. Homogenic events, such as
In contrast, heterogenic events, such as
One idea whose host population may accumulate members through both homogenesis and heterogenesis is the belief that the "head and shoulders" method of stock price forecasting is useless (Lynch, 2000). Many of the hosts of this idea received it (homogenically) from people who already held the belief. This makes the instances of their belief homoderivative. Yet some people also learn this idea "the hard way," by first learning a belief that they can forecast stock prices using a "head and shoulders" pattern, and then losing money trying to do so. In these people, the belief that one cannot forecast stock prices with the "head and shoulders" pattern is heteroderivative.
The concepts of replicator evolution and epidemiology directly pertain only to the homoderivative sector of an idea's host population. In this sector, we can properly refer to ideas as units of natural selection, similar to the neural instructions discussed by Cloak. This allows a very restricted denotation in which a thought contagion is defined as a homoderivative idea, or a homoderivative memory item. The definition of a particular thought contagion contains an abstract sameness criterion like the ones defining ideas, but also includes a criterion of causality, namely, that it is homoderivative. So each thought contagion has a corresponding idea: the idea defined without reference to homoderivative causation.
Although many instances of the belief that "head and shoulders" makes useless stock forecasting do not occur as a thought contagion, one can still understand virtually all of the idea's ongoing proliferation in terms of thought contagions. Simply divide the hosts into two groups: those who have learned the uselessness of "head and shoulders" from others and those who learned the idea through money-losing investments. The first group can be studied directly as thought contagion hosts. The second group's growth rate can be studied as a function of the frequency of the spread of the idea that "head and shoulders" is useful for stock forecasting. So the study of idea replication can be valuable to understanding the growth or decline of both of these constituent groups, whose combined numbers include nearly all the people who believe that "head and shoulders" is a useless stock forecasting method.
Some ideas are neither thought contagions nor thought contagion-derived in most of their instances. For these ideas, the study of thought contagions is of little use. Consider what happens when an earthquake of harmless but noticeable intensity strikes Los Angeles. Millions of people suddenly have the idea that an earthquake has struck on that particular day. So at first the host population does not result primarily from idea copying. In fact, the people who directly experience the quake may remain a majority of the idea's hosts, especially if the quake is too mild to get much news coverage. Replicator theory has extremely limited relevance in studying this kind of host population growth. The theory best applies to the limited but still vast realm of thought contagions and thought contagion-derived ideas.
The ability to unambiguously identify ideas as homoderivative now becomes
crucial to thought contagion theory. For instance, when someone receives
the hell idea from one person and the imminent doomsday idea from another,
is the resulting A*B combination homoderivative or heteroderivative? The
A*B host has copied both ideas from pre-existing hosts. But the event that
actually forms A*B, namely
The problem can be resolved by recalling that ideas replicate only with
respect to an abstraction. Although the A*B host is homoderivative with
respect to abstraction A, and homoderivative with respect to abstraction
B, the instance of A*B is clearly heteroderivative with respect to abstraction
A*B. So A*B formed by the event
Yet as mentioned earlier, A*B can also propagate as a set by the non-parental
conversion event
Disingenuous propagation raises similar questions. Consider the following ideas:
Idea P is the belief that "bee pollen invigorates."
Idea Q is awareness of the "bee pollen invigorates" proposition.
Idea R is the belief that "it is profitable for bee pollen merchants to tell customers that bee pollen invigorates."
If a merchant holds the ~P*Q*R combination, then a disingenuous propagation event might then be represented as
This event clearly involves replication with respect to the awareness
idea Q, and the profitability idea R. Yet this event alone is not homogenic
with respect to P, because the merchant is a non-host of P. Still, there
is a mechanism of indirect replication in this case. If the P*Q*R host
goes on to pay the merchant, it helps preserve the idea R in the merchant
and also pays helps pay the bills that the merchant needs paid in order
to continue instilling P*Q in new hosts. This form of proliferation is
already recognized in such fields as marketing science and consumer education,
and hence is not a distinctly evolutionary epidemiological insight. Also,
the actual profitability of transmitting P*Q involves numerous business
and marketing science considerations, such as how many people are persuadable,
how much they are willing to spend, how much the bee pollen costs to produce
and distribute, etc. Understanding the disingenuous spread of P*Q*R is
largely covered by older disciplines, so the propagation event here serves
mainly to illustrate how it is diagrammed and in what sense it can involve
replication of P. Note that the event
Simple non-commercial lies also qualify as events in which the transmitting
party holds an awareness of a proposition but not the belief in that proposition.
If the lie is believed by the recipient, it would take the form
Before proceeding, it helps to first give a formal definition of the word "replicator," as used in evolutionary cultural replicator theory and evolutionary epidemiology of ideas, and potentially in evolutionary replicator theory more broadly. The definition used here is as follows:
REPLICATOR: An thing whose instantiation depended critically on causation by prior instantiation of the same thing.
The "thing" (which can also be called an "item" or "entity") here is identified using the abstraction system being used by the observer or other investigator. Calling two things "the same" is taken to mean "of the same kind." Identification of two things as being "the same" or "of the same kind" is done only with respect to an abstraction system, or system of sameness criteria, of the an observer or investigator. (The section below on the fundamental role of abstraction in science goes into this further. We can equivalently define a replicator as "an instantiation of an abstraction that was caused by prior instantiation of the same abstraction," though such wording could confuse many readers, especially those who misread the word "abstraction" in this context as indicating detachment from reality.) The "thing" considered as a replicator can include the form taken by a molecule, as may be the case with prions. It can be a type of atomic nucleus, such as a short-lived excited form Uranium 236, whose prompt fission sends out neutrons that convert Uranium 235 into similarly unstable Uranium 236 nuclei. It can be the software of a computer virus. It can be a gene. It can be one of the elementary self-replicating instructions discussed in Cloak (1973). It can be a physical process, such as a candle flame generated by using a prior instantiation of "the same" process. It can be a pattern of such processes, as when a binary number is specified by candle flames lit or not lit as caused by a prior instantiation of "the same" pattern of processes.. It can be a quantum state, such as the spin state of a proton that has been made to have "the same" spin state of another proton as caused by the spin state of that other proton. These are just a few of the possibilities, as the focus here is on the evolutionary epidemiology of ideas. Note that the new instantiation of the item need not be entirely caused by prior instantiation of the same item. For example, the replication of a prion may depend not only on a pre-existing prion, but also upon the presence of water molecules, mineral ions, enzymes, and so on.
For a given thing, (self-replicating or not), one can usually make a long list of causal precursors -- some of them distant or very abstractly defined. For instance, one can (for what its worth) list the electron rest mass, charge conservation, etc. on a very long list of causal factors for some person's belief that Julius Caesar crossed the Rubicon (to choose a rather arbirary example). A defining issue in deciding whether something fits the definition of "replicator" or not is whether the potentially very long list of causal precursor factors includes at least one prior instance of the same thing. When the "thing" is the belief that Julius Caesar crossed the Rubicon, prior instantiation of that belief needs to occur in the list of causal precursor factors of that instance of the belief in order for that instance of the belief to be called a replicator. This minimalist definition of the term "replicator" allows for parsimony in evolutionary replicator theory. The theory amounts to causal analysis of items considered replicators and (potentially vast and varied) chain reactions of the associated causal events that qualify as replication events. (On generalizing the term "homoderivative" to apply to all sorts of things other than ideas, we could equivalently define "replicator" as synonymous with "homoderivative entity" or "homoderivative thing.")
The definition given above is somewhat different from the one given by Dawkins (1982), where he states:
I define a replicator as anything in the universe of which copies are made. (p. 83)
One difference is that the definition given in this paper renders more explicitly what it means for "copies" to be "made" of something. The making of a "copy" is explicitly defined in a minimalistic, hence parsimonious way as consisting of a prior instantiation of something playing a critical causal role in a subsequent instantiation of "the same" thing.
The definition given in this paper also requires only knowledge of past causation in order to ascertain the status of a thing (or item, entity) as a replicator. This can make it easier to apply, since one need not know if something eventually will be copied in order to decide if it is a replicator or not. Reference only to past causation may also allow for more inclusiveness: things caused by prior instantiations of "the same thing" are typically also among the phenomena one wishes to understand in terms of evolutionary replicator theory. The definition given in this paper can, in many cases depending upon the abstractions or sameness criteria in use, leave out things considered as being first in a chain (or lineage) of replicators. (In case it becomes important to include such cases, one can redefine the word replicator more inclusively to denote an entity whose instantiation depended critically on causation by prior instantiation of the same thing or that played a critical role in causing a later instantiation of the same thing.)
It is important to note that, with the definition of replicator used in this paper, the "sameness" of large numbers of entities does not itself indicate the presence of a replicator. In the earlier example where an earthquake causes millions of people to simultaneously form the belief that an earthquake has struck on given day, the initial wave of believers may include no instances of the belief that qualify as replicators. The definition of replicator used in this paper is not intended as a blanket label for all instances of all beliefs.
Thought Contagion, Formally Defined
A specific kind of replicator is the type defined below as a thought contagion, definition 1. Defining the term thought contagion technically without reference to other neologisms, we have:
THOUGHT CONTAGION: 1. A memory item, or portion of an organism's neurally-stored information, identified using the abstraction system of the observer, whose instantiation depended critically on causation by prior instantiation of the same memory item in one or more other organisms' nervous systems. ("Sameness" of memory items is determined with respect to the above-mentioned abstraction system of the observer.) 2. The process of repeated causation of new instantiations of memory items in which the causation of those new instantiations depends critically upon the prior instantiation of the same memory item in one or more other organisms' nervous systems.
The causation of a new instantiation can happen by a great many routes. The role a thought contagion plays in causing a new instantiation can seem rather passive, as when parents spend days trying to decide which word they will use for naming a newborn. Or the idea can play a strong role in manipulating host's communication behaviors toward retransmission, as when the idea of astrological compatibility motivates believers to retransmit to all potential mates. The process of causing a new instantiation can even be lengthy and complicated. For example, someone may spend years of trial and error efforts to impart idea A into a best friend. That friend may even rely on his or her own creativity during the process of becoming a new host of A. Yet if the idea transition depended critically upon the prior person having idea A, then all the talking, interaction, and even creative components are still summarized in the abstract form A + ~A --> 2A and counted as a single replication event for the idea A as a thought contagion.
The above definition may be seen as a formalization of the definition given in Lynch (1996), in which a thought contagion is simply defined as a self-propagating idea. Inasmuch as any idea can play at least a passive role in self-propagation by merely existing in someone's brain, [Lynch, 2001b], the more formal definition of "thought contagion" is consistent with the simpler 1996 definition. (Ideas that play additional active roles in self-propagation generally call for thought contagion analysis the most clearly.) Definition 1 of thought contagion is also equivalent to the terms homderivative idea and homoderivative memory item introduced earlier.
Although the above definition of thought contagion covers a vast range of phenomena, it is not intended to cover everything that one might want to analyze with evolutionary cultural replicator theory. Artifacts, co-dependencies, chain letters, groups, behaviors, certain propagating margin calls in financial markets, certain other financial chain reactions, institutions, entire societies, and numerous other social or cultural phenomena may be discussed using various kinds of replicator models. The strength of the replicator concept varies across such a wide range of phenomena. For those phenomena, different terms for the replicators involved might also be used. For example, the text of a paper or electronic chain letter can be called a text contagion, and studied as a type of cultural replicator. In cases where phrases are sent from recall by a person's brain, however, the text might best be considered a relic or a phenotype of a thought contagion. The latter usage of the term "phenotype" having been developed by Cloak [1973].
Many of the conceptual and analytical tools used to consider thought contagions also pertain to the various other kinds of cultural replicators, though often with modification. Still, the similarities allow for scientists to study a range of different kinds of cultural replicators. As with memory items, one still needs abstractions to consider two behaviors or two artifacts "the same." That means that one still needs to use abstractions to view behaviors or artifacts as replicators. For example, it takes an abstraction to say that two people both performed "the same" behavior of tying a shoe. One does not mean that hands followed the exact same trajectories over time, since no two people's hands are exactly identical. Some hands have fewer than five digits, others have extra digits, while still others have arthritic joints. Even for a single individual, the trajectories followed in tying a shoe can vary dramatically, while the physiological state of the hands and body also change from moment to moment and year to year. What one calls "the behavior" of tying a shoe is recognized more for the outcome of a secure bow made by one person than the specific behavior leading to that result. People also learn different ways of tying a shoe: single-handedly or left-handedly, while others learn to tie bows that have slight topological differences such as a clockwise or counterclockwise twist. Compounding the diversity is the huge variety of shoes and shoe laces. A very young child needs a certain amount of experiential learning just to know that a shoe-tying behavior is happening. Cultural replication can only happen with respect to an abstraction, whether it involves internal memory items or external behaviors and artifacts or any combination of them.
While a behavior such as tying a shoe is not itself a thought contagion, the knowledge of how to tie a shoe with a bow does spread as a thought contagion. The ease of untying a bow would have given people incentive to learn the knowledge of how to tie a bow from other people seen doing it. This is immitation in the classic sense. As the knowledge of how to tie a shoe with a bow spread in adults, parents had motives for inculcating the knowledge into their children: doing so saved them from having to dig terrible knots out of their children's shoelaces or from dealing with shoes falling off of their children or from always having to help the children with their shoes.
When artifacts are viewed as replicators, most involve brains at some point in the causal pathway to forming new "copies." Bows tied in shoelaces do not replicate themselves, and they are usually not even used for clues about how to tie shoelaces. Instead, bows result from behaviors that result from stored knowledge, with the behaviors playing an essential role in communicating the knowledge from brain to brain. The bow as an artifact helps call attention to the fact that there is a useful bow-tying behavior and knowledge of how to tie a bow. The artifacts of bows, the behavior of tying bows, and the knowledge of how to tie a bow all depend upon each other for propagation. They all depend on the persistence of memory for preservation, as the bows and the behaviors of tying them usually remain dormant while people sleep with their shoes off and untied.
A computer virus, however, can be seen as an artifactual replicator that usually spreads from machine to machine (or disk to disk, etc.) without being hosted by a new brain for every replication. The instantiations of the computer virus are not thought contagions, but the algorithm of the virus can be a thought contagion: the prankster who initially wrote the code may find his code being learned and copied by other pranksters. We thus have artifactual and brain-stored replicators. Similarly chain letters that manipulate recipients into retransmitting can be viewed as artifactual replicators. Especially in cases where the retransmission can be done by clicking a few buttons on a computer, replication may proceed without the recipient even reading the whole letter or knowing just what message it contains. The program trading of securities, currencies, and commodities can also result in chain reactions of propagating trading orders.
Similarly, one can investigate the role an enzyme plays in causing new instantiations of "the same" enzyme in one or many cells. Such investigation can find that nucleic acids usually if not always play a crucial ("central") role in causing new "copies" of the enzymea fact that has centered the analysis upon the nucleic acids rather than the enzymes as replicators. But the enzyme can still be viewed as a very indirect, multistage replicator. Cytochrome C, for instance, helps produce ATP that in turn helps produce new "copies" of cytochrome C. Yet much as one does not say that a protein enzyme is or contains a "gene," so too does one not say that an artifact is or contains a thought contagion. Still, an artifact can function as a medium of communicating knowledge of how to make new copies of "the same" artifact, and this difference from the role of proteins in biology suggests that there should not be a "central dogma" in the evolutionary epidemiology of ideas.
The various preliminary distinctions that go into defining the term "thought contagion" may appear to be another example of how the evolutionary epidemiology of ideas departs from analogy to genetics. Yet chemistry also has many distinctions that place nucleic acids in a very small subclass. Molecules can be catalysts or not. They can also be autocatalysts or heterocatalysts. The early oceans may have contained oligonucleotides formed by polymerization of smaller components. Some would have formed by non-autocatalytic polymerization, even in outer space. But if any did behave autocatalytically, they would have proliferated into predominance. They would have been a subclass of molecules generally, much as thought contagions are a special subclass of memory items.
Another distinction worth noting is that the term "thought contagion," by referring to a memory item in an individual, is a term intended more for the analysis of social micro-contagion processes than social macro-contagion processes. Examples of micro-contagion include the person-to-person spread of beliefs about the stocks of particular companies [e.g., Lynch, (2000), (2001a); Shiller, (2000)] as well as broader economic ideas such as the belief that "the economy is worsening." An example of macro-contagion is when recession in an entire country helps cause recession in other countries that are the first country's main trading partners.
In human societies, not all social contagion processes fall neatly into the micro or macro categories. Social contagion may be a mixture of macro-contagion and micro-contagion, as when one country's recession contributes to causing recession in another country both by way of gross trading volume (macro) and by way of person-to-person ideas about recession spreading across boarders. The latter can involve centralized or de-centralized communication, topics discussed again in section 10. Centrally magnified micro-contagion can also happen, as when newscasters and news writers spread ideas about recessionary times to each other by way of the very media output they produce, while at the same time that mass media output spreads the ideas to thousands or millions of times more people in the general public. We also have massively mutual micro-contagion, as when a large crowd chanting a slogan grows by gaining the attention of nearby individuals. When such demonstrations gain media attention, they can give rise to centralized massively mutual micro-contagion, and perhaps also to macro-contagion events that spawn similar demonstrations in different people. We even have intermediate-scale social contagion that would could be seen as falling between macro- and micro- levels. An example might be chanting crowds of small to moderate size achieving enough attention to spread both the chant slogan and the idea of chanting in small crowds to other people who may form into new little chanting crowds of their own.
There are no perfect analogies to physical or biological contagion processes, but there are perhaps some pedagogically useful comparisons. One way to understand the distinctions is by considering what happens in star formation. New stars blow away surrounding gases with their radiation pressure, stellar winds, local heating, and eventually with novas and supernovas. That compresses the surrounding gasses and helps new stars to form, leading to a grand chain reaction running on an astronomical scale until it depletes gasses to below a star-forming threshold. This may be viewed as a physical macro-contagion or macro-replication process. At the same time, there is a micro-level process of nuclear chain reactions involving the fusion of hydrogen in the stars. With nuclear fusion, the process is actually massively mutual, since one fusion event generally does not lead directly to another. Instead, many fusion events produce the overall heat needed to cause many more fusion events.
Much as micro- and macro- replication processes are not in themselves sufficient to understand the many kinds of observed phenomena in the universe, so too are social micro- and macro- processes not the only forces acting in societies. Moreover, simple replication does not itself amount to evolution by natural selection either in nuclear, stellar, chemical, biological, or social phenomena. Hence the need for further discussion, even in a paper that focuses mainly on evolutionary cultural micro-contagion.
Definition 2 of the term thought contagion refers to the process of propagation of the memory items identified in definition 1. Definition 2 is thus an abstract noun compared to definition 1.
Within the present theoretical framework for analyzing the process of
thought contagion and evolutionary cultural replication, replication of
ideas and contagion of ideas each have the same formal and functional definitions
in terms of one instance of an idea causing a subsequent instance of the
same idea. Cultural replicator theory of ideas is thus also called contagion
psychology of ideas, and evolutionary cultural replicator theory of
ideas is also called evolutionary contagion psychology of ideas.
[The term contagion psychology was previously used in Lynch (1999),
and possibly before that.] Since things other than ideas may be analyzed
as replicators or contagions external to the brain, contagion psychology
and evolutionary contagion psychology can include more than internally-stored
ideas. Internal ideas may be seen as holding a special place mainly because
of their role in forming decisions. Contagion psychology and evolutionary
contagion psychology, in turn, fall within the scope of
population psychology.
Cloak's paper on elementary self-replicating instructions includes a concept of the "size" of the replicators, inasmuch as "elementary" can be construed to mean "of minimal size" within some kind of metric. More specifically, Cloak lists three criteria that need to be met in order for an instruction to qualify as an "elementary self-replicating instruction" or SRI. To quote, "An SRI is an instruction which (1) engages in events of self-replication, (2) engages in behavioral events of at least one other sort, and (3) is not composed of parts which are SRIs; i.e., which meet criteria (1) and (2). Criterion (3) is, of course, what makes an SRI 'elementary.'"
Determination of "elementary" status, that is, criterion (3), is a fairly easy thing to measure for segments of DNA or their corresponding proteins, as well as for pieces of text. There, a common system of quantization between replicas may simplify the task of determining whether or not another self-replicating instruction can be within a given one, rendering the latter one non-elementary. For DNA, all the candidates under consideration as elementary self-replicating instructions are composed of a finite number of nucleotide subunits. There are thus, only a finite (though potentially very large) number of subsets of nucleotides that could be combined to form the one being tested for elementary status. This renders the task of checking whether any of those subsets of nucleotides are themselves self-replicating a finite task, even if the combinatorics requires a vast number of possibilities to be checked. Likewise for strings of letters forming a piece of text: there are only a finite, even if huge, number of subsets of the letters of the text. Yet with ideas, beliefs and other information stored in the brain, we must currently do without generally accepted methods of quantizing the information into minimal constituent parts that are the same across different instantiations of an idea, belief, or other piece of information. Instead, we must consider the possibility that the mode of storage differs across different brains, as if different organisms used different genetic materials or each individual copy of a text were written in its own unique language and unique alphabet. Despite this difference in storage, we generally still agree that two people can have "the same" piece of information stored in their brains. For example, two people can each know the same phone number for the telephone company. Though the number itself is a piece of digital information, its storage details may differ across different people. Likewise, many people around the world know the first 6 digits of the number pi, and have the information stored in their brains. Digital information can also be stored in a vast number of ways in computers, using options such as the processes of transistor current flows, ferromagnetic materials, electrostatic charges, laser markings, etc., but with beliefs stored in brains, there need not always be consensus about whether units akin to digits can be found, let alone what those units are. Nevertheless, there are ways of partially determining whether one idea may be considered as a component of another.
The most we can really say is that a set of ideas is, in general, a partially ordered set, as defined in set theory [note5]. One rather arbitrary system of partial ordering is based on the size of conjunctions of the ideas identified by a particular abstraction system. Take the ideas expressed by the following 4 statements, for instance:
Idea A: "There is only one true God."
Idea B: "Christ is Lord."
Idea C: "Unbelievers are damned."
Idea D: "Earthly life is better among believers."
We have no basis for saying if A > B, A < C, etc. That is, we have no basis of saying if A is larger than B in an ordering system, or if A is smaller than C in that ordering system.
We can, however, say A*B > A, and A*B > B.
But you cannot say A*B > C or A*B < B*C, etc.
Still, we can say A*B*C > A*B, A*B*C > A*C, etc.
In other words, if the hypothetical faith only says "There is only one true God," and "Christ is Lord," we cannot, for instance, say that it is "bigger" (has more "size") than the faith that only says "Christ is Lord" and "Unbelievers are damned." Nor can we compare the "sizes" of these component beliefs. We also do not know whether or not the individual ideas A, B, C, and D can be composed of other ideas, or if so, whether there is a unique or preferred way of doing so. Each statement can, for instance, be considered as incorporating ideas about language, about what a God is, about what a true God is, etc.
For completely unordered sets such as S
In special cases, the most elementary self-replicating self-replicating
instruction of a set of self-replicating instructions may nevertheless
be identified, as with the set of ideas S
When it is possible to construct a set of ideas that is fully ordered,
the full ordering of that set may become a useful empirical consideration,
as with the set of ideas
Even outside of fully ordered sets, it is still possible to describe
some elements as "elementary" after Cloak [1973].
For example, if A, B, C, and D are each in some set U, where U also contains
various or even all possible combinations of A, B, C, and D, then U still
has no elements that are "smaller" (in our conjugation metric) than A,
B, C, and D. Therefore, U may be a partially ordered set, and still have
a unique subset of "elementary" elements. That subset may even include
all of elements from which other the rest of the elements of the set U
are derived by conjunction. The number of "elementary" elements in the
subset need not include all possibilities, as in the case where
U = {A, A*B
Cloak [1973] recognized that one could not simply proceed in discussing elementary self-replicating instructions without defining what it means for an instruction to be "elementary." The present section elaborates and clarifies that subject in terms of set theory and metric spaces. Later works, such as Dawkins [1976] simply jump into discussion of "a unit of cultural transmission" or "a unit of imitation" without defining what it is for some item of culture or imitation to constitute a "unit," which presumably means something that is the smallest possible of its kind. To be smallest possible of a kind requires one to explicitly state the system of measure with respect to which an item is smallest of its kind, or more precisely, the smallest element of a set. That is, some kind of a metric space must be defined, even if it is only based upon numbers of distinct items joined in combination. The widespread lack of definition of this aspect of "cultural units" or "units of imitation" in works on evolutionary cultural replicator theory following Dawkins (1976) has probably contributed to a perception that the definability of "units" idea was nothing more than an unstated premise evaded by most of the literature.
Massively Cooperative Propagation
Though religious idea sets such as those above spread by inducing a
wide variety of one-to-one transmission events, some can also induce events
involving large numbers of participants. In multistage events, there can
also be large numbers of participants in some stages and few in others.
Thus, (retaining the symbol meanings of the preceding section) an individual
might spread A*B*C through the event
Now consider what happens when adding a few more ideas:
Idea E: "Love your neighbor as yourself"
Idea F: "The Spirit lives in the community of believers"
Idea G: "Church X is God's Church."
As noted in Lynch [1996], ideas that cause greater cooperation among their own hosts can thereby achieve propagation advantages by raising prosperity and suppressing the sort of personal conflicts that may lead to dropouts. The "love your neighbor" idea E even motivates more one-on-one evangelism in some settings while promoting collective idea propagation in other settings. The idea expands the range of unbelievers that adherents wish to save from damnation: instead of just wanting to "save" a few friends and relatives, they want to "save" even strangers. Yet the idea also works in many ways to improve community life among believers, also noted in Lynch [1996] and documented by Stark for early Christians in The Rise of Christianity [Stark, 1996]. So when an individual Christian attempts to spread ideas A through G to a pagan, the pagan may well remain unconvinced. Yet the pagan realizes that idea D is testable merely by spending some time among believers, and without waiting for an afterlife. In ancient times, they would have found a Christian community thriving from cooperation, and taking better care of each other's misfortunes than did the pagans. That would have impressed them that at least the idea D part of the Christian message was true. This in turn would have made it cognitively easier to believe the "Spirit" idea F, and the "Spirit" idea F would make it easier to believe the "God's Church" idea G. Finally, that would make it easier to believe other Christian tenets such as ideas A, B, and C.
In addition to these cooperative conversion effects, a community can
also achieve greater rates of preventing the dropout event
For some ideas, the effect on rates of cooperative propagation is particularly
striking. With Hutterites (denoted here in simplified form with idea symbol
H), for instance, we observe many events of the form
The phenomenon of large numbers of humans propagating a movement by acting in concert has been cited as evidence for the group selection of human genes. Wilson and Sober [1994], for instance, take the colony behavior of Hutterites as evidence of innate hive-like social dispositions arising from group-selected genes. Yet the ideological imperatives discussed above can explain the evolution of a Hutterite belief set in terms of the evolutionary epidemiology of ideas alonewithout requiring any new innate factors beyond a very general capacity to learn and comply with cultural mores. Likewise, the Christian propagation advantages that arise through community behavior do not suggest a special genetic mechanism. As important as they are, massively cooperative idea replication events remain a subclass of idea replication events in general. The fact that they happen for a variety of ideas in various populations suggests that communities can achieve certain idea transmission effects beyond the capacities of single individuals. Yet it does not of itself show that the ideas must be relying on an innate imperative for highly collective action.
Centralized idea transmission stands in contrast to massively cooperative
propagation. Here, a very few people act to spread an idea to vast numbers
by events such as
Mass media often play an important role in centralized idea replication events. When they do, most of the recipients cannot go on to play the role of idea retransmitter by the same kind of centralized event. This may at times make centralized communication less suitable for study with replicator theory. It can also at times limit the role of distributed replication: much as humans can acquire legionella bacteria centrally from central air conditioning systems, so too can we acquire ideas by way of intentionally central idea spreading systems. Still, contagious transmission can happen through centralized communication, as discussed in section 7 above.
Another feature of centralized communication is that not all of the people transmitting the message are actually hosts of the idea. The idea that "Coke is the real thing," for instance, has probably been broadcast by many celebrities who prefer to drink something entirely different in the privacy of their homes. This again limits the usefulness of replication considerations, as sender and receiver do not always have "the same" idea. The sender, for instance, may only have the idea that it is profitable to say "Coke is the real thing" rather than actually believing in the message. Yet as with non-centralized profitable communications, people who adopt the idea often spend money that pays merchants and advertisers to send the message again and again. This is a phenomenon recognized and applied long before the advent of evolutionary cultural replicator theory, and already studied in marketing science.
Mature recipients of centralized messages often understand that the sender may not believe what she says, thus limiting receptivity to the message. Communications from friends and family generally do not have an apparent profit motive, too, and can thus enjoy a more privileged reception. Desires to maintain harmonious relationships also cause people to pay closer attention to messages from direct acquaintances. This helps maintain the importance of non-centralized communications from immediate family and acquaintances in the age of mass media. In turn, it prevents all of idea propagation and change from being reducible to existing work in media studies or marketing science.
Centralized communication can range from honest, candid, and helpful to deceptive, manipulative, and cynically engineered. Much of it is appropriately called propaganda. Since the most central communicator(s) or designers of propaganda often do not believe what they wish to persuade others to believe, the distinctions drawn in section 6 for non-centralized lies still apply.
By focusing on the evolutionary aspects of mass belief, thought contagion analysis is not intended to replace existing understandings of what happens in propaganda. It does, however, posit that there is much more to the formation of mass opinion than the purely centralized engineering of beliefs. Yet it does not deny that, for instance, centrally generated plans to repeat a big lie loudly and often do influence public opinion. Indeed, science itself has increasingly become a venue for propaganda, as can be discerned from titles of papers. If a paper that is centrally published in a journal has a title that reads "Why the evolution hoax is harming biology," one can best understand the title as a piece of propaganda -- especially if the article does nothing to establish that evolution is a hoax and only uses the word "hoax" in its title. Though titles of similar form may sometimes arise from poor word choices, this particular title suggests to the reader that evolution is a hoax, that everyone already knows that evolution is a hoax, that evolution is harming biology, that the only real question to be answered is why, and that this question is already widely asked by biologists. Its promulgation as propaganda arises from the fact that a title is much more widely read than a full article. While the title used in this example was made up for purposes of illustration, papers with titles of similar manipulation and deception structure have been published in various disciplines, including disciplines concerned with the study of culture. Another form of propaganda is to use resume inflation to lend credibility to otherwise dubious claims. This has recently become a greater problem in science due to the ease with which online identities can be manufactured, "borrowed," or stolen. Thought contagion analysis does contribute to the understanding of the propaganda of mass movements or incipient mass movements by helping to explain what leads to some of the intense ideologies whose adherents often generate propaganda. Thought contagion analysis also helps explain why "rank and file" adherents work to disseminate the centrally-produced propaganda of some movements more than others. This is particularly useful in cases when ideological variations formed from propaganda campaigns start to take on evolutionary contagion trajectories of their own. Moreover, propaganda often exploits pre-existing ideologies and values whose evolution and prevalence can also be largely understood in terms of thought contagion. Finally, thought contagion analysis can help understand the spread of propaganda tactics among propagandists who work for various and often competing causes.
The continued importance of distributed communications even manifests itself in the contagious propagation of ideas specifying which centralized sources to consume. The Bible, for instance, may look like a centralized communication, coming from mass production plants ever since moveable type was invented. But the idea of buying and reading it propagates person to person in the population, along with various ideas specifying what passages to emphasize and what they mean. Another example discussed in Lynch [1996] was the localized propagation of ideas for listening to centralized political talk-radio shows. Listeners to shows with pro-business, anti-tax messages play the radio openly in the workplace, allowing co-workers to learn about the show and its message. But listeners to shows that might politically offend the management feel inclined to use headphones, which blocks the local retransmission of their political and show-preferences. Even that most centralized of modern media, television, exhibits natural selection processes, as when the remote control caused a trend toward greater violence, sex, and melodramathe types of content that happen to stop the passing "channel surfer." [note7] Far from replacing distributed ideological evolution, the centralized media have instead opened up new realms for propagation and natural selection in competing idea systems.
Communication can also be both centralized and massively cooperative. An example of massively cooperative centralized communication is when protest rallies in one city generate media attention that leads to similar protest rallies in other cities. It typically takes many protesters together to gain the centralized media attention. But once they do gain centralized media attention their message and the idea of staging a protest rally to communicate that message are transmitted to many more people, often in distant cities.
The Fundamental Role of Abstraction in Science
The great potential for confusion in discussing cultural phenomena forces explicit reference to systems of abstraction. Yet the axiom of abstraction, fundamental to all of science, is generally invoked on a tacit level in most disciplines. Indeed, the axiom of abstraction is also known as the axiom of comprehension. It merely states that for any attribute, one can define a set of all the entities that have that attribute. Saying that two entities are "the same," then, is tacit shorthand for saying that they are elements of the same set, whose inclusion criterion is the attribute. The phrase "the same with respect to an abstraction" serves to remind readers of the underlying mental operations involved in calling any two entities "the same."
In the physical sciences, abstraction manifests itself every time someone refers to an entity as having the attribute of being a "water molecule." "Water molecule," is an abstraction which scientists have found useful for discussing phenomena. What they really mean by "water molecule" is a particular pattern of matter and energy. It is a way of calling two or more patterns of matter and energy "the same," ignoring such differences as location, velocity, rotational states, vibrational states, oxygen 18, Hydrogen 2, nuclear spin states, electron spin states, electron excitation states, quark states, etc.
Is it ever legitimate to identify patterns amid all this seething matter and energy? A philosophy of science question. Yet the grouping of vast categories of objects into sets defined by attributes seems to be a fundamental feature of human thought. Without formally justifying the practice, one can note that this can be an efficient way of storing vast amounts of knowledge in a finite brain. One can also theorize that evolution has favored brains that do this because it provides sufficiently functional representations of reality to give an animal a survival and reproductive advantage. But these arguments themselves depend on abstractions.
Regardless of why we rely so much on abstractions, science as we know it is largely a project to develop and test ever stronger systems of abstractions with which to describe and comprehend reality, often calling them "theoretical constructs.". The "stronger" systems of abstractions are defined as those which explain more of reality with reliance on fewer essential abstractions. Yet deep questions remain on whether nature will some day indulge the physicist's quest for the kinds of "ultimate" abstractions of "grand unification theories." Will our desire for ever more unifying abstractions instead be frustrated by some fundamental complexities of a reality not designed specifically to please the human mind? This we have yet to learn.
Despite advances in fundamental knowledge, we cannot run through the equations of quantum mechanics to explain something as complicated as a cell or a DNA molecule. So we invoke further layers of abstractions for these vastly complicated systems. Instead of representing "adenine" in terms of a chemical formula, a single letter is used. And "little things" like the dissociation of H+ from phosphate groups and particular serpentine shape traced out by the molecule are ignored. Compared to the level of elementary particles, molecular biologists focus on patterns of patterns of patterns, layers of abstraction removed from what physicists call "fundamental."
A great deal of science went into demonstrating that "genetic material" is a useful abstraction, and that "nucleic acid" can be used as an equivalent abstraction here on Earth. Likewise for the discovery that "nucleotides" are useful abstractions for describing nucleic acids. Yet the pre-molecular concept of "gene" was a useful abstraction, and its very utility provided impetus to the research that led to more powerful molecular abstractions. More recent arguments over different molecular definitions of "the gene" amount to arguments about which abstractions are stronger. Yet the "the gene" started as an abstraction based on studies of entire organisms, and has progressed into an abstraction based on studies of molecules.
Materials can also be manipulated to fit a set of desired abstractions, as has been done with digital electronics. One voltage interval is chosen to represent the abstraction "1," and another the abstraction "0," with all sorts of "low level noise" to be ignored. Analog vacuum tubes, transistors, and other devices are then wired to produce abstractly bimodal outputs corresponding to abstract operations on abstractly bimodal inputs. Hence the "NOR gate," for instance, which produces a "low voltage" output whenever either of the inputs falls in the range arbitrarily labeled as "high" in voltage, even though it is almost always less than 5 volts in the early 21st century.
What is called a "1" or a "0" by engineers is actually a process of flowing charge carriers in most of these circuits. Special circuits called "static memories" are actually composed of these logic gates or transistors with special feedback connections. Thus, a "1" or a "0" in static RAM, for instance, is still an abstractly defined process for most current technologies. A set of "analog" transistors are wired together so as to behave in a relatively bimodal fashion--a fashion corresponding to abstractions ("1" and "0") used by the engineer who designed the circuit. Then, when the process of currents running through the transistors is initiated from a power supply set in a certain range of voltages, currents and voltages in ranges that the engineer labeled as "0" or "1" are produced. The persistence, or "sameness" (with respect to abstraction) of a "0" or "1" from one millisecond or microsecond to the next is then maintained by the dynamic process of currents flowing. "Static" RAM is thus based on a dynamic process. Nevertheless, the theoretical construct, or abstraction of "static 1" or "static 0" is useful in describing and analyzing the behavior of the device. Likewise, terms such as "memory content" are useful theoretical constructs and abstractions. On a more fundamental level, the theoretical constructs of "charge carriers" such as "holes" are useful in the design of semiconductors.
Still higher levels of abstraction arise when referring to "software," where specific voltages and even patterns of "1's" and "0's" can be ignored in deciding whether two "programs" are "the same." Science again builds layers of abstractions, this time working from the low level up rather than the high level down.
Because "replication" depends on "sameness," and the "sameness" of any two "water molecules," "DNA molecules," etc. exists only with respect to an abstraction, it follows that replication only happens with respect to an abstraction. Abstraction systems and sameness criteria are important in all evolutionary replicator theories. Yet "replication" also involves the notion of causation. Specifically, the word "replicator," can be replaced with the cumbersome phrase "instantiation of an abstraction that causes a new instantiation of the same abstraction." This is the tacit meaning in use for all kinds of evolutionary replicator theories ranging from the "artificial life" to thought contagions to autocatalytic molecules that might evolve in the oceans of Jupiter's moon Europa.
Thought contagions are abstractions currently based on macroscopic observation. The extent to which it will stay at this level remains to be seen. In any case, the systems of abstractions used to study them do not require that they be "detected" by some kind of molecular or microscopic means any more than software science requires that "programs" be "detected" by visual of chemical tests. Many social scientists already treat "Nazism" or "monotheism" as "real" phenomena without providing any molecular or cellular basis. Many even go on to attempt to get others to hold "the same" idea as theirs about various social science theories. In doing so, they unwittingly affirm the underlying premise of thought contagion theory, that one person can cause another person to have "the same" idea by way of "communication." They need only admit further that different ideas can repeatedly spread at different rates before seeing that the evolutionary epidemiology of ideas can follow mathematically. There too, many of them affirm a principle of evolutionary epidemiology of ideas by avidly trying to make their own beliefs spread at a greater rate than competing beliefs. This is not the same thing as accepting specific thought contagion hypotheses, but it does show tacit acceptance for the underlying tenets of the theoretical framework of evolutionary epidemiology of ideas.
Nevertheless, some may experience discomfort from the fact that science has not discovered an absolute, fundamental, or privileged system of abstractions with which to discuss socially transmitted information. But the "sameness" of any two people's learned information about a topic depends on the abstraction system of the observer. The "units" in evolutionary epidemiology of ideas are no more fundamental than are meters, inches, and so forth for measuring distance or location. With genes, nature has rendered a few very similar abstraction systems far more powerful than the alternatives, so that scientists do not notice the role of the observer's abstraction systems. But there are many ways to define "anti-abortionism," for instancewhich is another way of saying that the abstraction system and survey questions can be constructed in many different ways.
Evolutionary epidemiology of ideas will probably have to do without the "fundamental unit" idea much as physicists have now abandoned the idea of an absolute coordinate system (framework of location and time abstractions) for the universe. Not only are meters and seconds arbitrary units of measure, but measurement results depend on the framework in which measurement is taken. More fundamentally, just as physicists now accept that the "simultaneity of events" exists only with respect to an observer's coordinate system, so too must thought contagionists accept that the "sameness of ideas" exists only with respect to an observer's abstraction system. While this is not a "special theory of cultural relativity," it does show scientific precedent for admitting that multiple systems of abstraction can apply to a given class of phenomena. It does not mean that all abstraction systems are equally useful, but it does suggest that science can proceed without the assumption that there must be one abstraction system that is universally right for the evolutionary epidemiology of ideas.
Evolution is fundamentally a quantitative as well as a qualitative theory. The abstractions that allow us to discuss ideas and idea replication events set the stage for discussing quantitative event rates. Differences between ideas in the rates of replication and termination events are what give rise to the natural selection of ideas. They are hence essential to the evolution by natural selection of ideas. From the fact that event rate parameters differ between ideas, it follows mathematically that natural selection must happen. Yet this statement can be rendered much more specific. The mathematically necessary consequences of differing event rate parameters can be characterized, using systems of differential equations for an ideas' host populations versus time. Defining the units and quantifying the events thus allows us to analyze the host population dynamics in ideological evolution.
The particular terms needed for a system of equations to model the population dynamics of a specific set of ideas will depend upon what kinds of events occur at non-negligible rates for the ideas being modeled. A vast range of event forms can be conceived, but the rarest of them do not need to be modeled in order to gain acceptable accuracymuch as the rarest side reactions in a chemical process can be ignored in quantitative chemistry. Models will in general have to account for at least two population groups, namely, those for hosts and non-hosts of an idea. Accounting for additional population groups is accomplished by expansion to a system of more than two equations. Most models will also have account for some very common types of replication events, such as simple parent to child transmission, non-offspring conversion, spontaneous dropout, and mortality. Two-equation systems to model these basic kinds of events therefore constitute a starting point for mathematical modeling of differential idea transmission, allowing further elaborations to be added as required.
The following two differential equations pertain to two ideas whose
host populations are represented by
The other parameters in the equations have the following meanings: t is time in years, a is host age, p is the age of a second personthe idea propagatorused in places where two people's ages are involved.
R1(a) is the fertility rate for idea-1, in
children per host of age a per yearthe quantity parental parameter.
a1 is the fraction per year of idea-1 hosts who convert to idea-2 without any prior idea-2 hosts teaching them, or who at least convert by mechanisms that do not depend on the size of the idea-2 host population. Preventing such "dropouts" is one form of preservational advantage (information longevity advantage) for idea-1. Finally, M1(a) is the rate of mortality per age a idea-1 host per year.
Swapping "2" subscripts for "1" subscripts in the above sentences gives
the parameter definitions corresponding to changes in idea-2 prevalence.
[note8]
(1)
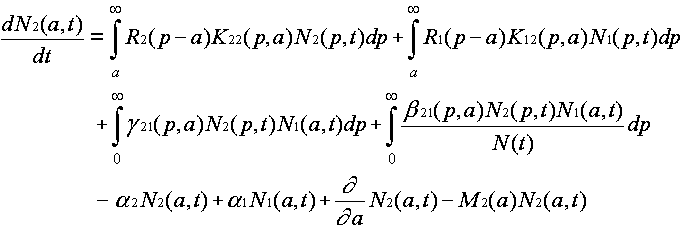 (2)
(2)
The first two terms in equation 1 are the parental terms. In the first
term, the group of
In practice, there may be many cases where a useful mathematical model
can be attained by treating the parent to child idea transmission as if
it all happened when the children reached the single age c1,
the average age at which they pick up idea-1 from a parent. Also,
one might find that the transmission rate per child depends very little
on parent age differences within the mainstream host child raising years.
If this is true, then one can replace the
The next two terms in the equation concern the non-parental conversion
mode of transmission. The first of these is the one that is sensitive to
the total number of potential converts,
The following two terms (beginning of line 3) express the "spontaneous" dropout rate for idea-1 and idea-2, respectively. "Spontaneous" dropout rates are assumed to be proportional simply to the number of hosts capable of dropping out. Such rates could also be modeled as age-dependent rates by replacing a1 with a1(a) and a2 with a2(a).
The next term is the partial derivative of
The final term expresses the mortality rate as a function of age among idea-1 hosts. Mortality per host per year at age a (a kind of actuarial data) is simply multiplied by the number of hosts at age a to give the overall rate.
Equation 2 above models the same kinds of propagation processes for idea-2 as are modeled for idea-1. Equations 1 and 2 form a system of non-linear differential equations modeling the interdependent propagation of idea-1 and idea-2.
What follows are five more equations that go with equations 1 and 2,
defining relationships between the propagation parameters. Equation 3 states
that all offspring of idea-1 parents end up holding either idea-1 or idea-2.
Equation 4 says the same thing for idea-2 parents. Equations 5 and 6 state
that one group's non-parental conversion gains are the other group's non-parental
conversion losses, so that the net non-parental gain to the whole population
is 0. The last equation merely defines the function
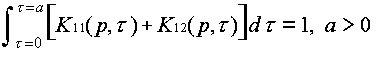 a>0 (3)
a>0 (3)
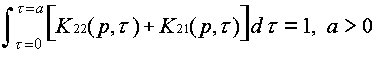 a>0 (4)
a>0 (4)
![]() (5)
(5)
![]() (6)
(6)
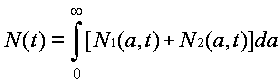 (7)
(7)
Not all idea propagation events are given their own separate terms in equations 1 and 2. For instance, if it frequently happened that idea-1 hosts produced idea-2 offspring who then converted their parents to idea-2, then the rate at which idea-1 hosts were non-parentally converted to idea-2 might depend greatly on how many children they had. A new term might have to be added to the equations to make this phenomenon adequately modeled. The model can, in fact, be made arbitrarily complex, but it is obviously desirable to keep it as simple as the application permits.
Another elaboration of the equations, and one that might interest mathematical sociobiologists, is the explicit inclusion of specific genes and their propagation parameters into the picture. Such a model would analyze "host populations" of ideas, genes, and idea-gene combinations all in the same system of equations. Of course, one sets the a's, b's, and g's to zero wherever the propagation of a gene is modeled. The parental terms become more numerous and take on a diversity of forms corresponding to all the combinations of genes and ideas that can occur in two mating adults. Keeping track of the homozygous and heterozygous combinations would also cause more population parameters and equations. Each possible outcome of each parent combination must have its own term in one of the equations. Such equations embody no a priori assertions about the relative importance of either genetic change or cultural change over a modeled time span, nor any assertions of how strongly or weakly prior genetic and cultural evolution constrains the course of change over that time span. Instead, they allow for considering these matters on a case by case basis once the requisite starting data are fed into the models.
Although emotional and cognitive receptivity factors are not readily conspicuous in equations 1 and 2, they are in fact represented. The reason is the K 's, b's, and g's are measures of successful idea transfer events. As such, they are composites of both the rates at which propagation is attempted and the rates at which it is cognitively and emotionally well-received. Likewise, the "spontaneous" dropout rates (a's) include their own products of cognition and emotion.
Equations 1 and 2 model idea transmission using aggregate propagation parameters. One way of elaborating on the equations is to use stratified propagation parameters. This may be especially useful for peer-to-peer transmission. Thus, one may wish to replace the g's with more elaborate functions that could then model situations where there are vast differences between the number of people persuaded by the best communicators and the number persuaded by the median communicator. The best communicators could be those who exhibit high levels of transmissivity or whose messages enjoy high levels of receptivity in recipients, or both. For example, they could be the people who have access to centralized communication. Or they could be authority figures, people whose messages carry special weight to thousands or millions of others. Often, authority figures are given more access to centralized communication, and those who happen to have access to centralized communication are ipso facto considered authority figures. While such stratified communication is common, its effect can be reduced in a few ways. First, the influence of a host population's best communicators may in many cases be achieved at the expense of more ordinary (e.g., median) communicators, so that the aggregate propagation parameters are approximately conserved. Second, the host populations of two competing ideas may exhibit similar patterns of stratified communication, which could lead the aggregate values of propagation parameters to predominate in overall idea transmission. Moreover, for certain important personal ideas, messages from one's immediate friends and family carry much more weight than do messages from centralized communicators or distant authority figures. This still leaves ideas for which stratified propagation parameters have a major influence on transmission as a function of time. For example, if, for some interval of communication rates, the communication rates are inversely proportional to the fraction of people having higher communication rates without merely concentrating communication done by lesser communicators, then a strongly self-accelerating transmission can result. In such a case, communicators at the 90th percentile would have 5 times the media communication rate. Those at the 99th percentile would have 50 times the median communication rate. Those at the 99.9th percentile would have 500 times the median communication rate, and so forth. Those at the level of a top one in a million communicator would have 500,000 times the median communication rate. As the propagating idea accumulates its first 10 hosts, its first 100 hosts, its first 1000 hosts and so forth, its early growth, or growth through the first several orders of magnitude from a host population of 1 could thus proceed at a faster than exponential rate. Increasing amounts of idea propagation would result from communication by grand disseminators. There are many other ways that communication parameters can be stratified besides inverse proportionality to the fraction of people having higher communication rates. An entire separate paper could be written to model stratified propagation parameters and their effects on idea transmission as a function of time.
Stratified communication can also matter in another way: messages from grand disseminators may affect median communication rates, or the communication rates of "ordinary" communicators. For example, when early peer-to-peer transmission of a new idea causes it to spread into its first authority figure, most of the rest of the idea's hosts may become aware that their idea is now being expressed by an authority figure. That can make "ordinary" and median communicators feel more confident and willing to express the idea. That can raise average and median levels of transmissivity for the idea. The original idea can also spread in combination with the added idea that an authority figure is endorsing it. This can raise the receptivity of people who are newly exposed to the original idea or who are re-exposed to the original idea plus the added idea of endorsement by an authority figure. Many people are more interested in hearing an idea if they believe that it is also expressed by an authority figure. Many are also more inclined to adopt an idea if they believe it is also expressed by an authority figure. In fact, evolution can even cause beliefs that were not originally expressed by an authority figure to become widely misattributed to an authority figure. Such evolution is briefly discussed in Lynch, 2001a. In short, the idea as misattributed can simply out-propagate the same idea with the initial or source communicator correctly identified. All of these mechanisms provide additional potential causes for self-accelerating propagation of ideas, and can again lead to faster than exponential propagation. They can also greatly affect the maximum prevalence of an idea -- perhaps by many orders of magnitude in extreme cases. In particular, an idea that has very low prevalence can become far more prevalent when it is adopted and retransmitted by a first one or more authority figures. However, when two competing ideas have both achieved substantial propagation, they can both enjoy similar effects from grand disseminators and the influence of grand disseminators on communications between "ordinary" peers. The measurement of aggregate propagation parameters in such populations can also take into account the effects of grand disseminators and the influence they have on communication among "ordinary" (e.g., median) peers. Still, for some kinds of ideas, the effects from grand disseminators and their influence on communication rates of "ordinary" communicators should be explicitly considered when modeling the transmission of an idea through its early orders of magnitude of growth from a host population of 1. These effects, which are also not modeled in equations 1 and 2 above, would again require at least another full paper to consider.
Anti-competitor propagation advantage is also not fully represented by the K 's, b's, and g's of equations 1 and 2. Part of the reason is that this mode can occur in quite a wide range of ways. It makes a big difference, for instance, whether the idea-1 group merely bans idea-2 proselytizing or launches a idea-2 extermination campaign. Moreover, the effectiveness of such measures does not vary as a simple function of the idea host populations. The Nazis, for instance, became dramatically more harmful to competitors after they became numerous enough to gain political power. Such phenomena may well defy mathematical modeling techniques based solely on predicting host populations versus time, and require the detailed modeling of political processes as well.
The K's, b's, and g's each modeled as overall effective rates of idea transmission. The K's, for instance, do not indicate how many times a parent needs to repeat a message to her children before it is effectively learned. The b's, and g's likewise do not reflect how many a message was voiced from hosts to a non-host before that non-host converted. A more detailed model might therefore break down these parameters into the subfactors of transmissivity, a measure of how often each host attempts to transmit an idea, and receptivity, a measure of the likelihood each host to non-host transmission attempt has of actually imparting the idea to a new person. Much research has been done on how various components of receptivity affect the diffusion rates of innovations Rogers [1983]. Receptivity parameters can also might in turn be broken down still further to reflect different probabilities of idea acceptance on first, second, third, etc. exposure. Modeling idea-based differences in receptivity to idea-1 or idea-2 that result from the presence or absence of third or fourth (etc.) ideas requires expansion to more than two main equations, but may be necessary in cases where the empirical evidence suggests that non-host populations have both "susceptible" and "non-susceptible" subsectors. All of these possible elaborations generate more complicated mathematics.
All idea propagation involves some level of transmissivity, receptivity, and longevity of the ideas. Evolution among ideas arises from differences in transmissivity, receptivity, and longevity. [note9] One of the differences between the evolutionary epidemiology of ideas and "classical" social science is that the evolutionary epidemiology of ideas looks at all three of these general factors, while the classical social sciences often neglect transmissivity and longevity in favor of receptivity. Though social science approaches other than the evolutionary epidemiology of ideas do acknowledge transmissivity differences in mass media, questions of why a whole society came to believe something are often posed as questions of "why did the people want to believe." Investigating such questions does provide useful information, but not a complete picture.
Equations 1 and 2 model fairly ideal cases of the two-idea propagation problem, but also serve as the kernel of a multi-equation system for cases involving three or more ideas with host populations N1, N2, N3, etc. The equations offer a sample of the kinds of terms that can arise in realistic applications, and they illustrate that a unified quantitative analysis can be given to qualitatively dissimilar modes of propagation. They also illustrate the concept that once the main mechanisms of an idea's propagation have been empirically discovered and measured, the degree to which each mechanism contributes to instantaneous rates of propagation can be mathematically modeled. The resulting systems of differential equations govern host populations as a function of time, and so can be used to generate limited predictions of what will happen if the equation parameters remain reasonably constant in non-chaotic intervals. It must be noted, however, that any validity the predictions may have can be diminished or eliminated by giving the predictions to the population whose ideological and behavioral future is being predicted. Generally, the more widely disseminated such predictions are in the population whose future is being predicted, the less likly it is that the predictions will match reality, up to the point where the predictions are no better than random guesses. This is because giving a prediction to the population whose future is being predicted alters the beliefs and behavior of that very population. If one tries to give a higher order prediction that includes the presence of the first order prediction, then that too will alter the population's ideas and behavior. Likewise for third order, forth order, fifth order, etc. predictions. The resulting infinite regress can make it impossible to both publicize a prediction in the population whose behavior is being predicted and at the same time have that prediction be convincing and reliable. Disseminated predictions can of course be self-fulfilling, but they can also be self-refuting and well as various other possibilities. Similar problems may be encountered by economists and financial analysts who publicize their predictions, even when their jobs demand them to do so.
The model presented above treats each idea as being either instantiated
or not instantiated in a given person. An alternative is to model ideas
as a continuum, in which there are an infinite number of mutually exclusive
degrees of "hostship" that are instantiated or not. In other words, each
individual is considered to occupy a single point on an ideological continuum.
Strictly speaking, it would no longer model "replication." Instead, each
point on the continuum would exhibit its own birth and mortality rate.
Parents at each degree of the trait would have a distribution of rates
of imparting all possible degrees of the trait to offspring. People communicating
with non-offspring would have distributions of second-party transfer rates
denoting how often they move people from one level of the trait to any
other level of the trait. Yet in measuring such continuum traits, investigators
generally end up assigning subjects to a finite number of trait levels
such as "strongly agree," "agree," "neutral," "disagree," and "strongly
disagree," designated here as ideas A, B, C, D, and E, respectively. This
returns us to dividing the total population into five (or alternately,
seven, ten, etc.) idea host populations with a corresponding number of
equations. Still, horizontal communication can involve heterogenic events
in which, for instance, someone in the "strongly agree" category gets someone
in the "disagree" category to transfer to the "neutral" category. As noted
earlier, not all ideas are replicators with respect to the abstraction
system in use, and an abstraction system using finitely stratified categories
would include both replication and non-replication (heterogenic) events.
However, certain unidirectional multistage heterogenic events can add up
to form homogenic (replication) events with multiple participants. If,
for instance, four distinct E-hosts convert an A-host into an E-host
with in stages (a separate E-host participating in each stage), then the
heterogenic stages still add up to a homogenic event. The first three events
taken individually are heterogenic:
Returning to non-stratified ideas, a great many phenomena that may look
like two-idea problems are in fact applications calling for three or more
equations. This includes cases that enumerate not only the hosts of an
idea, but also the exposed and unexposed non-hosts. For instance, if we
define idea P as belief in proposition X, then non-belief is ~P.
If
awareness of proposition X is designated as Q, then unawareness
is ~Q. An exposed non-host of the proposition is designated as ~P*Q. Exposed
hosts are P*Q. Unexposed non-hosts are ~P*~Q. The P*~Q combination presumably
has a host population of zero. The resulting mathematical model therefore
has three equations modeling
Host populations modeled as distributions in age and time in equations
1 and 2 can also be measured and modeled using a variety of additional
parameters such as those of geography (e.g., longitude and latitude), duration
of hostship (as distinct from the age of the host), family income, years
of schooling, etc. The R's, K's, b's,
g's,
a's,
and M's could be modeled with the additional parameters as well.
The R's, K's, b's,
g's,
a's,
and M's would then quantify a range of events that include individuals
changing not only their belief their host status, but also individuals
changing their locations, incomes, etc. The corresponding event diagrams
would include non-replication attributes, such as location coordinates
x, y, and z, so that the simple migration of an idea P*Q host from
location 1 to location 2 would become P
For certain classes of idea replication phenomena, one may wish to use still more elaborate or different kinds of event diagrams to explicitly demarcate the sex and mating status of idea hosts. This is particularly true for ideas that spread at very different rates among the sexes, such as the valuation of ones own chastity, preferences for long finger nails, the valuation of dangerous sports, preferences for wearing neckties, etc. It is also true of ideas that affect rates of marriage and divorce, which could include beliefs that marriage is sacred or beliefs that divorce is sinful.
One reasonable method of demarcating sex is to put a subscript "M" or "F" on letters designating ideas or idea conjunctions. Thus, in a female, idea A would be AF, while AM would represent the same idea in a male host. For conjunctions, parentheses would be added, so that A*B*C in a female would is represented as (A*B*C)F and in A*B*C a male is represented as (A*B*C)M.
A "Ä" symbol can then be used to represent the mating of two idea hosts. The formation of a mated couple can be represented by a mating event such as:
(A*B*C)F + (A*B*C)M
This reads "Female host of A*B*C plus male host of A*B*C yields an A*B*C couple."
Idea discordant matings can also be represented, as with
(A*B*C)F + (~A*B*~C)M
This reads "A female A*B*C host mates with a male ~A*B*~C host.
Divorce events can be essentially the reverse of mating events. If our idea-discordant couple divorces with no interim conversions, the event would be represented as:
(A*B*C)FÄ(~A*B*~C)M
Idea replication events can, of course, happen during a mated relationship. When one partner converts the mate, it is necessary to add another subscript to indicate who did the transmitting. Otherwise, the event could be confused with a spontaneous (self) conversion by one partner. Thus, we have, for example,
(A*B*C)FtÄ(~A*B*~C)M
The subscript "t" in (A*B*C)Ft indicates that the female is acting as idea transmitter in converting the male from ~A to A status. If a third party, such as a single male A*B*~C host does the converting, then the event is represented as:
(A*B*~C)Mt + (A*B*C)FÄ(~A*B*~C)M
If an A*B*C concordant couple has a female child and then inculcates her with A*B*C, then the event is represented in the multi-stage fashion as:
(A*B*C)FÄ(A*B*C)M
In the more abbreviated notation, this event is written as:
2(A*B*C)
Although this abbreviated form contains less information about the sex and mating status of participants, it has the advantage of showing the event's contribution to host populations more conspicuously: starting with two A*B*C hosts and ending up with three A*B*C hosts.
In principle, any number of people can be represented as mated in 3, 4, 5, etc. person relationships. Such combinations may need to be diagrammed when modeling the spread of polygamy ideas, for instance. Yet in many cultures, such relationships may be sufficiently uncommon for purposes of parent to child idea transmission that they can be neglected. Nevertheless, a reproduction event in a union of 3 or more should include a subscript r to indicate which two parties were the biological parents.
Still other symbols besides the "Ä" may be introduced to indicate other kinds of relationships besides mating. Tacitly, mathematical equations or computer simulations must keep track of certain relationships already, such as parent-child relationships. Parent-child relationships are already tacitly tracked in the equations given in the Units, Events, and Dynamics paper, which enumerate parent-child inculcations happening years after a childs birth. Computerized individual-based models would also keep track of who is parent to whom, even if no new symbol is introduced to represent the parent-child relationships the way "Ä" indicates mating relationships.
When modeling only monogamous male-female matings, enumerating the various combinations of ideas, sexes, and relationships is still a very complicated task. This may make analysis with systems of equations prohibitively difficult. Computer simulations with multi-agent models are, however, a promising way to proceed in quantitative studies of sex-linked ideas and idea-discordant mating.
With all of these event diagram elaborations to consider, as well as the difficulty of working with systems of complex equations, one can also use quantified propagation mechanisms to run predictive computer simulations of ideological evolution without the intermediate step of writing down differential equations. The latter method can run particularly well on systems based on generalized cellular automata, where each "cell" simulates the idea state and behavior of a person. Such an individual-based computer simulation is now feasible using the SWARM program developed by the Santa Fe Institute [Langton, 1998]. Another major advantage of such simulation is that it can take account of specific patters of mixing and contact between hosts and non-hosts, or more generally, between members of populations 1, 2, 3, etc. -- matters that can be extremely difficult and awkward to incorporate into a system of differential equations.
Equations 1 and 2 belong to a class of equations seldom, if ever, used in other fields: systems of non-linear, partial differentio-integral difference equations. As difference equations, they are further set apart by integration over a difference parameter. The unusual nature of the equations argues against the reliance upon metaphor for more than pedagogic or initial creative analogies in the evolutionary epidemiology of ideas. It also argues against trying to avoid or minimize the mathematics by grafting equations in from other fields. A similar case could be made for the study of replication phenomena on a planet where organic polymers with specific compositions and chirality features are found to catalyze the formation of inorganic crystals with specific compositions and chirality features, and where the specific types of crystals in turn catalyze those specific types of polymers. Ultimately, a formal analysis must be shown to stand on its own rather than left to rely upon metaphors.
The equations embody some of what is distinct about the epidemiology of ideas. Equations 1 and 2 could, in principle, be applied in the future to some unrelated phenomenon, but this would be an instance of mathematical thought contagion theory being used as a source or basis of metaphor, rather than an analysis based on metaphor to previously analyzed phenomena. That said, it is important to note that equations 1 and 2 compose just a small part of the literature on mathematical modeling of social evolution. Boyd and Richerson (1985), Cavalli-Sforza and Feldman (1981), Hamblin (1973), Bartholomew (1982), Witten (1991), and others provide extensive applications of mathematical modeling to social and cultural evolution.
One could well ask why equations 1 and 2 are as complicated as they are. While it is possible for an author to use extremely abstract mathematical operators that defy empirical determination while bewildering all but the most mathematically sophisticated readers, equations 1 and 2 were not written for abstruseness or escape from real world data. Indeed, all of the mathematical operations performed in the equations can be understood by any reader who has learned and retained two to four semesters of calculus--not exactly graduate school mathematics. Still, the equations are unusually elaborate, leading to the general question of whether or not that is really necessary. The answer is that complexity arises from the complexity of the social phenomena they are modeling. For instance, equations 1 and 2 unify the influences of peer to peer and parent to child transmission in a continuous model that does not impose arbitrary generations on the population. The first 4 terms in each equation are particularly elaborate, but they can handle such phenomena as the progression of a religion or worldview from mainly peer-to-peer transmission at low prevalence to mainly parent-to-child transmission at high prevalence. One misconceptions about thought contagions is that peer to peer transmission should always be faster than parent to child transmission. At its fastest, peer to peer transmission is in fact much faster. But with large, complex belief systems, peer to peer transmission takes a great deal of time and effort and may also encounter resistance due to cognitive and emotional factors, or competing ideological systems. As a movement wins more converts, it also starts to exhaust its supply of persuadable non-converts, forcing it to slow down even more. (Persuadability corresponds to "receptivity," a subfactor of the beta and gamma parameters of equations 1 and 2.)
Available data bear this out: Gallup polls have found that evangelical Christianity had a 34% prevalence in the USA during August 27-30, 1976, and a 44% of a 32% larger adult USA population 21.83 years later during June 22-23, 1998. That works out to a geometric mean growth rate of 2.541% per year, or one above-replacement convert made per adherent per 39.4 years. And that rate is a combination of parental and non-parental transmission, with believers typically opposing abortion, homosexuality, and other non-procreative behaviors. It also includes growth from increased life expectancy. Some of that results in more converts by or of senior citizens--but some of the growth also comes from a non-conversion increase in population due to long-time adherents saying alive longer. Even if we assume that the 1976 figure was high by 5% of the adult USA population and the 1998 figure was low by 5% of the adult USA population we still get only a roughly 3.802% geometric mean growth rate in prevalence, or one above-replacement convert made per adherent per 26.2 years. Either way, the growth rate of the whole movement is comparable to growth rates for some mainly parentally learned faiths. Thus, any equations that model the phenomenon must model parental and non-parental transmission modes simultaneously. Such a unified analysis of apples and oranges is what leads to great complexity in the equations. One bonus, however, is that the same equations apply during early phases of rapid peer to peer spread (abundant persuadable non-converts) and later phases of mainly parental transmission that occur after most persuadable people are already converted. This includes what might be called slow horizontal transmission: after exhausting the supply of persuadable non-adherents in a given decade, the movement must in effect wait for old unpersuadables to die off while new non-adherents grow up and pass into phases of relatively higher persuadability decades later. Even improved communication technology might make just a slight difference in this process: the main effect could be to accelerate the depletion of persuadable non-adherents without eliminating the long wait for new ones to grow up.
Such complex mixes of parental transmission, non-parental transmission, and mortality lead to rather elaborate systems of equations. In fact, the system of equations 1and 2 can even be expanded to deal with different levels of persuadability, as by dividing into populations N1, N2, N3, etc. corresponding to multiple ideas with various levels of persuadability (or persuasion immunity) to each other. The equations could actually have been written in a more complex but general form with summation (sigma) signs, subscripts i and j, as well as vertical and horizontal ellipses on top of all the integrals and derivatives. Instead, they were written in their more readable 2-equation kernel form.
Quantitative Analysis for Artifactual and Behavioral Abstractions
Although labels of individuals based solely on artifacts and externally visible or audible behaviors are not treated as ideas, or thought contagions in this paper, equations 1 and 2 can be generalized to apply to such labels. That is, equations 1 and 2 can model the natural selection of artifactual abstractions and external behavior abstractions as well as memory abstractions, provided that the individual person is still taken as the measure of transmission. For instance, N1 can be defined as the population possessing a certain kind of artifact, and N2 defined as the population not possessing that kind of artifact. Alternatively, N1 can be defined as the population exhibiting a certain external (e.g., motor) behavior, and N2 defined as the population not exhibiting that behavior. However, as population equations, equations 1 and 2 cannot directly model the number of copies of an artifact or the number of instances of an external behavior. The latter quantities would require modification of the equations or the development of different types of equations. One possible method of proceeding is to separately model the number of artifact copies or external behavior instances per person counted as "possessing" the artifact or exhibiting the behavior, and multiply this quantity by the numbers N1 or N2 as modeled by equations 1 and 2. Naturally, that is a much easier task in those cases where the behavior or artifact count per person can be treated as constant over the time interval under study. Such mathematical methods do not work, however, for entities that are said to be "the same" replicator whether instantiated in behavior, artifact, or brain. The replicated subset of combined behavioral, artifactual, and neurally stored information already goes by the term "culture," and so is not given additional nomenclature within this paper. The term "thought contagion" is used in this paper to refer only to the replicated subset of neurally-stored information, a subset of culture that thus receives identification.
The description of the evolutionary epidemiology of ideas as being about how ideas influence behaviors that propagate ideas need not be taken as implying a rigid idea/behavior dichotomy. The phenomenon we call "life" is a material process, and processes can be viewed as behaviors. Some behaviors can be labeled "internal" while others can be labeled "external" while still others can be labeled as mixes of "internal" and "external." The neural memory of anything is actually a process, as is the "static RAM" storage of a "1" or a "0" mentioned earlier. The axons and dendrites that form a synapse, for instance, are dynamic, ever-changing, metabolizing parts of cells. Their lipids, water, ions, proteins, etc. are all in states of flux at various rates. It is only through process that they remain "the same" (with respect to an abstraction) from one day to the next, or one year to the next. Thus, the concept of "memory item" or "idea" can be rephrased in terms of behaviors causing behaviors. The "internal" neural behaviors called "ideas" in the preceding sections can thus affect "external" speech behavior, for instance. That "external" behavior can then affect the "internal" behavior of another person in such a way as to cause a new "internal" behavior that is "the same" (with respect to an abstraction, or theoretical construct) as the "internal" behavior of the first person.
Such theoretical constructs handle the recurrence of external behaviors in a single organism as well. They are also intended to remain consistent with recent and potential future observations of internal behaviors using PET scans, microelectrodes, etc. -- the neurobiological research.
In behavioral terminology, a phenomenon or experience that people might identify as an "idea" held for (say) 20 years can be considered an ongoing 20 year internal behavior, and identified by abstraction as is the case with more temporary external behaviors such as shoe tying discussed in section 7. A focus on behaviors need not be limited to the macroscopic actions of such body parts as muscles, bones, and skin, but may also include the microscopic actions of neurons and the overall actions of such internal organs as the brain. (There is no magical barrier that allows for consideration of behavior only of body parts and organs other than the brain and nervous system.)
While some may prefer to focus exclusively on external behaviors in hopes of attaining reliable observability, the goal of observability is not always met. External behaviors can be deceptive, a fact routinely demonstrated by magicians but also evidenced in such areas as the unreliability of eye witnesses. There are also many important phenomena poorly handled by considering only external behaviors without reference to internal behaviors or ideas. For example, a specific set of ideas about how to hijack a plane and use it to attack a skyscraper can be replicated among a number of people even if the actual behavior of attacking a skyscraper with a hijacked plane has never before taken place. People involved in such a plot can all have "the same" idea (i.e., instantiate the same internal behavior abstraction) long before the actual attack. To understand the causes of such an attack, one simply must pay attention to ideas or abstractions about internal behaviors in people before the first such attack ever happens. This includes paying attention to whole systems of belief as well as ideologically-motivated attack plans. Many less spectacular events also call for explanation in terms of ideas or abstractions about internal behaviors.
Among the less spectacular phenomena that call for explanations in terms of ideas or abstractions are urban legends. To understand such stories, one must consider how much attention they command in the people who have learned them. A vivid story that causes people to keep thinking about it and considering its implications can, by that fact alone, achieve more retellings per host. If a vivid story provokes 10 minutes of thought from its average host in a given week, while a bland story provokes only 1 minute of thought from its average host in a given week, the difference in time spent thinking about the two stories can easily cause a large difference in how many times hosts of the two stories repeat the stories to new listeners. One must consider internal behaviors involving internal memories of the stories in order to make such an analysis.
In financial markets, one frequently must consider not only the securities, cash, merchandise, and documents that people hold, but also what beliefs they hold about companies. Consideration of both artifacts and beliefs, then, allows one to explain why a company such as eToys once had a very high share price even as it was loosing money and heading for collapse: shareholder beliefs about the company's prospects were out of line with reality. Again, internal beliefs cannot be ignored to perform the analysis (Lynch, 2000).
Despite the importance of ideas and internal behaviors in human culture and society, belief systems that oversimplify culture and society can proliferate in part because of the greater ease of generating, transmitting, and remembering simple theories. For example, theories that all the interesting aspects of culture can be understood in terms of genetic replicators alone are easier to create, learn, and spread to others than are theories that analyze both genes and ideas as replicators. Likewise, theories asserting that the only true cultural replicators consist of external behaviors and artifacts (to the exclusion of ideas and internal behaviors) may also be easier to create, learn, and spread than theories that allow for a wider range of cultural replicators along with genetic replicators. Since ideas and beliefs are more personal, some people also have adverse emotional reactions to the analysis of ideas and beliefs as replicators, even if they are more comfortable with the analysis of less personal things such as genes, behaviors, and artifacts. Still, no matter how simple or emotionally unthreatening it is, a theory must be comprehensive enough to account for the full range of observations. For that purpose, a very general theory of evolution that recognizes replicating genes, ideas, behaviors, artifacts, etc. may be necessary. Yet the aspects of such a theory that deal with ideas as replicators must, as in this paper, address complex issues such as the identification of the replicators involved. Methods of analyzing ideas as replicators without falling into conceptual chaos may thus be less simple, and hence harder to create, learn, and spread than are corresponding methods for analyzing genes, artifacts, or external behaviors as replicators. However, the greater explanatory, predictive, and practical utility of a more comprehensive theory may justify the extra effort needed for studying it.
Qualitative and Quantitative Evolution: Population Creativity
As mentioned earlier, occurrence rates of heterogenic events often depend on the prevalence of precursor ideas. When vigorous precursor ideas proliferate, they achieve substantial odds of causing the creation of ideas that only a rare host can form. Additionally, when the new idea is simply defined as a combination of two or more precursor ideas, the vigorous propagation of the precursors greatly hastens the arrival of the combination. So the independent spread of ideas W, X and Y hastens the formation of the combination W*X*Y. Indeed, if W, X and Y spread vigorously, W*X*Y can pop up explosively among many widely separated individuals. A novel combination such as W*X*Y may have interesting psychological or cultural effects in its own right, forming one source of qualitative evolution or innovation resulting from quantitative evolution.
Yet if W*X*Y tends to inspire hosts to create idea Z, then Z, too can pop up explosively among widely separated individuals. This much acclaimed feature in the innovation of ideas shows that creativity is largely a population phenomenon. As an idealized and simplified illustration, suppose that W, X, and Y have simultaneously spread exponentially at the same time constant from a prevalence of one per billion people to a prevalence of one per 10,000 people, and that they are randomly mixed in the population such that the prevalence of W*X*Y is equal to the product of the separate prevalences of W, X, and Y. This can be stated as P(W*X*Y) = P(W)*P(X)*P(Y). The prevalences P(W), P(X), and P(Y) have each risen steadily by 5 orders of magnitude since their inception. Yet P(W*X*Y) is still only 10-12, making W*X*Y unlikely to exist in any human being at all. Now suppose that each one grows at the same rate for just one more order of magnitude to a prevalence of one in 1000. Now P(W*X*Y) = P(W)*P(X)*P(Y) = 10-9. This transition would only have taken one sixth of the propagation history of W, X, and Y, yet during that last span of time, W*X*Y goes from being unlikely to exist in any humans at all to having an expectation value of six hosts in a population of six billion humans. The six hosts are simply the 6 x 109 humans multiplied by the new P(W*X*Y) = P(W)*P(X)*P(Y) = 10-9; that is, 6 x 109 x 10-9 = 6 humans. We would therefore expect that the first 6 hosts of W*X*Y all occur during the last 1/6 of the transmission history of W, X, and Y by the time those six occurrences of W*X*Y come into being. If the formation of idea Z depends upon having W*X*Y, then it will become appreciably likely only during the last 1/6 of the transmission history of W, X, and Y. If, additionally, only 0.1% of hosts of W*X*Y have enough free time, creativity, motivation, etc. to actually form Z, then the likelihood of Z arising when X*Y*Z has six hosts is still only about 0.6%. However, when W, X, and Y have spread by still another order of magnitude to a prevalence of 1% each, we should expect to see the first 6 formations of Z as well. We therefore expect the first 6 formations of Z to all happen in a span of just 1/7 of the propagation history of W, X, and Y. Moreover, by this time, W, X, and Y are all within a few orders of magnitude of the maximum potential propagation of 100% of the population. If they each level off abruptly at 10% of the population, then the first occurrences of Z would all happen in the last 1/4 (= 2/8) of the propagation of W, X, and Y. We should also expect such temporal clustering to be even more pronounced in cases where still more precursor ideas are involved, such as when ideas T, U, V, W, X, and Y are all needed as precursors in a single host before Z is formed. In that case, the first 6 occurrences of T*U*V*W*X*Y would not happen until T, U, V, W, X, and Y had all reached a prevalence of 3.2%, which does not happen until they are all in the last 1/16 of their propagation history. If we again assume that only 0.1% of T*U*V*W*X*Y hosts have enough free time, creativity, motivation, etc. to actually form Z, then the first 6 independently formed instances of Z do not arise until T, U, V, W, X, and Y are at the very end of their transmission history (to the idealized, abrupt 10% prevalence limit). The arithmetic works out such that first one instance of Z is not expected to occur until T, U, V, W, X, and Y reach the last 1.62% of their propagation histories! Hence, all independently formed instances of Z that happen while T, U, V, W, X, and Y are still spreading happen during the last 1.62 percent of the propagation histories of T, U, V, W, X, and Y, clearly a case of strong temporal clustering! Complex feats of creativity may thus exhibit very intense temporal clustering. Also, the present illustrations assumes perfectly random mixing of the precursor ideas in the total population. In the event that the precursor ideas are geographically concentrated, then they may propagate for some time in isolation before reaching each other at all, and then suddenly arrive at each other's geographical frontiers. This could cause still more intense temporal clustering of both the combining of precursors in individual hosts and the formation of the new idea.
By adding assumptions, estimates, or measurements of how often Z is formed by hosts of T*U*V*W*X*Y, as well as how quickly Z spreads once formed, one may also investigate or model how many times Z is independently formed before such a high fraction of T*U*V*W*X*Y hosts are aware of prior formations of Z that they do not endeavor to re-invent Z themselves. Such an analysis could include an estimate or measurement of how long after achieving the T*U*V*W*X*Y combination it takes a creator of Z to actually complete the creation of Z. It might, for example, take 5 years of holding T*U*V*W*X*Y in order for the average independent creator of Z to actually create Z. During that time, the earliest formations of Z may be spreading through the population and reducing the number of T*U*V*W*X*Y hosts who are giving any effort to creating Z. If Z is a scientific idea, for instance, it may take decades to spread through the scientific community after its first formation. But if the first formation depended upon having thousands or millions of T*U*V*W*X*Y hosts, then Z would continue to be formed independently even as the earliest occurrences are spreading [note11]. Eventually, however, the prevalence of T*U*V*W*X*Y*~Z falls below the level at which we expect more than one host of T*U*V*W*X*Y to form Z in a lifetime. The spread of replicated Z thus attenuates the rate of independent creation of Z. However, people seeking to independently form new ideas can reasonably expect to discover that they have company, at least in some of the ideas that they independently form. Relatively prolific creativity might therefore be a characteristic common amongst those who actually achieve "priority," or first host status in some of the new ideas that they independently form.
Although the analysis here does elaborate somewhat upon the similar analysis of "Qualitative and Quantitative Evolution" in section ten of Lynch (1991), it is still handled on a simplified basis mainly intended to illustrate a proposed principle. A more detailed analysis could be made using equations 1 and 2. An intermediate analysis could also be done by assuming that mortality, dropout, and birth rates are zero, that everyone is of the same age, and that peer-to-peer persuasions are not influenced by special closed relationships such as spouses or siblings. That would reduce equations 1 and 2 to logistic equations, allowing logistic curves ("s-curves") [e.g., Hamblin, 1973] to replace exponentials that reach an abrupt ceiling. Such "s-curves" are also empirically well documented by Rogers [1983] for the diffusion of innovations. The more refined analysis of new formation rates of Z in terms of separate s-curves for the propagation of T, U, V, W, X, and Y would deserve a separate paper.
The analysis above also simplifies the relative timing by which T, U, V, W, X, and Y reach wide prevalence in the population. In reality, the separate ideas can begin spreading at different times and spread at different rates. Thus, independent temporally clustered formations of Z should be expected to occur for many innovations, but not for all innovations. Those where it is expected are the ones where multiple precursor ideas are all spreading through orders of magnitude of prevalence during the same time interval. For example, if 6 precursor ideas T, U, V, W, X, and Y are involved, the effect could still be observed if T reaches 50% prevalence while U reaches 0.1% prevalence and V reaches 10% prevalence and W reaches 1% prevalence and X reaches 4% prevalence and Y reaches 25% prevalence. As those percentages were reached, the prevalence of the combination T*U*V*W*X*Y (assuming random propagation into combination) be P(T)*P(U)*P(V)*P(W)*P(X)*P(Y) = 5 x 10-9. At that level, 30 people (out of a human population of 6 billion) would have the combination T*U*V*W*X*Y. If 1/10 of those people independently form Z, then there would be 3 independent formations of Z that would tend to be temporally clustered toward the time when T, U, V, W, X, Y were reaching 50%, 0.1%, 10%, 1%, 4%, and 25%, respectively. The degree of temporal clustering depends upon the exact values of P(T), P(U), P(V), P(W), P(X), and P(Y) as functions of time, a matter which should be analyzed formally in a separate paper. Such an analysis would show that the P(T), P(U), P(V), P(W), P(X), and P(Y) need not all increase at the same rate (e.g., with similar time constants) in order to produce the effect. It would, however, show that similar propagation rates is one of the sufficient conditions that produce strong temporal clustering of the independent formation of Z.
While the above argument offers a theoretical explanation for the temporal clustering of inventions within a population of people who do not live in cultural isolation from each other, previous work by Murdoch [1956] finds that high rates of parallel invention happen in "peoples of the same or related cultures," but only rarely "among unconnected peoples of differing culture." Murdoch [1956] even attributes this to differing rates of synthesis, which corresponds to the more formal explanation of ideas propagating into separate but equivalent combinations as given above. Murdoch's findings of parallel invention in "peoples of the same or related cultures" are thus consistent with the above analysis of temporal clustering, as are his observations and analyses of the difference in rates of parallel invention in related versus unconnected cultures.
In special cases, there may be one or more combinations of elements of the set Q = {T, U, V, W, X, Y} for which the propagation rate of the combined form exceeds the propagation rate for the individual ideas T, U, V, W, X, Y. In this case, the first occurrence of the smaller combination can happen much sooner than the first occurrence of T*U*V*W*X*Y that would result from independent propagation of the ideas T, U, V, W, X, and Y. In general, it is more difficult to learn, retain, and retransmit a combination of ideas than it is to learn, retain, and retransmit the ideas of that combination one at a time. However, there are also cases when something about the nature of a combination of ideas causes adherents to make the attempt more often, or to be more preoccupied with the ideas, etc. Those cases allow for a selection-enhanced and selection-accelerated first occurrence of T*U*V*W*X*Y, and potentially Z as well. In very special cases, there may be a two-idea combination of the ideas in Q that is more vigorous than the one-idea propagation rates of its constituents, then a 3-idea combination formed by adding just one of the 4 remaining ideas to the two-idea combination that spreads still more vigorously, a 4-idea combination that spreads more vigorously by addition of one of the remaining 3 ideas to the 3-idea combination, a 5-idea combination that spreads more vigorously by addition of one of the remaining 2 ideas to the 4-idea combination, and finally, the 6-idea combination spreading still more vigorously than that 5-idea combination. This produces a greatly accelerated evolutionary pathway to forming T*U*V*W*X*Y, and eventually to forming the new idea Z from T*U*V*W*X*Y. In such cases, the separate prevalences of T, U, V, W, X, and Y is no longer as stringent a determining factor in how long it takes to form T*U*V*W*X*Y or Z. We should also expect much less tendency for the first several independent formations of Z to be temporally clustered. The argument is general, so that one could set Q' = {I1, I2, ... Im}, and find, in special cases, that there exists a collection of simultaneous conditions such that propagation rate increases for some sequence of combinations (Ii*Ij), (Ii*Ij*Ik), . . . (Ii*Ij*Ik*... In), . . . (Ii*Ij*Ik*... Ip) that would produce the combination (Ii*Ij*Ik*... Ip) at a greatly accelerated pace, with i, j, k, and n all being unequal integers less than integer p, p being less than or equal to integer m, and each member of the sequence of combinations having all the elements Q' that the previous one has plus one more added. (For p < 5, the one, two, or three unneeded subscript numbers n, k, j would be dropped. More subscript numbers would be used for n > 5, as indicated by the ellipsis, in which case a sub-subscript notation could be used to avoid the limited supply of letters.) The hastened first occurrence of Ii*Ij*Ik*... Ip would also greatly accelerate the first arrival of any ideas that are far more likely to be formed in those holding the combination Ii*Ij*Ik*... Ip than in people lacking that particular combination of precursor ideas. In the most special cases of all, it would be true that for all integers n from n = 2 through n = m, with integer m > 1, any combination of n of the elements {I1, ... Im} spreads more vigorously than any combination of fewer than n of the elements of {I1, ... Im} or any single element of {I1, ... Im}. That would tend to produce the most accelerated first occurrence of the combination (I1*... *Im). One could then say that the "closer" the n-element combination is to (I1* ... *Im), the greater is its propagation advantage over all combinations containing fewer than n elements.
The fact that having one or more sequences of ascending levels of propagation rates for incrementally larger combinations are mathematically special cases, however, can leave us with many occasions on which some idea is formed simultaneously and independently in widely separated individuals. Even in the special cases of sequences of ascending levels of propagation rates, there is still some temporal clustering associated with the temporally accelerating spread of the last idea (Im) of the combination. Either way, the resultant temporal clustering may lead many people to perceive an illusion of some kind of collective mind or unconscious mind working to produce new combinations of ideas and especially new inventions of ideas almost simultaneously.
Although it is mathematically a "special case" for there to exist even one sequence of combinations (Ii*Ij), (Ii*Ij*Ik), . . . (Ii*Ij*Ik*... In), . . . (Ii*Ij*Ik*... Ip) as for which propagation rates increase with each idea added in combination, that does not mean that it is a rare case. The total number of such sequences is equal to m!, a number which increases very quickly with increasing number of elements m of set Q'. The probability that any randomly chosen m-member sequence of combinations has increasing propagation rates may, for instance, decline exponentially with m, but m! increases faster than an exponential. (Combinations that asymtotically approach upper limits of propagation via functions such as logistic curves or S-curves prevent the overall growth rate of larger and larger combinations from needing to be arbitrarily fast.) Although this does not take account of the limitations of short-term and long-term human, it may still result in more sequences of increasingly vigorous combinations existing for larger sets of ideas (larger values of m). The effect might therefore happen more often in the evolution of large systems of ideas or beliefs.
Some processes similar to the formation of new idea combinations may be involved in the arrival of polygenic biological innovations. The gradual accumulation of inherited characteristics by variation and natural selection has been explored in the biological literature ever since Darwin's 1859 book On The Origin of Species by Means of Natural Selection [Darwin, 1859], and is therefore only briefly discussed here. It does, however, bear such a conceptual resemblance to the sequential mode of producing the idea combination Ii*Ij*Ik*... Im that it has played a historical role in causing the formation of the various ideas about the evolution of ideas and of culture generally.
One possible difference is that in biological evolution, the mere propagation into combination of several genes in combination that had once spread in isolation from each other would be sufficient to cause a new polygenic trait to emerge. It is not necessary to assume that the propagation of once-separate genes into a new combination somehow causes the formation of a completely new gene, or to adjust the meaning of the word "gene" so as to call the new combination "a new gene." A new combination might in some instances, however, cause selection advantages for specific kinds of mutations in existing genes and for novel genes caused by other sources of variation, such as chromosome crossovers and the action of transposons. If a new combination of genes is adaptive, then evolution should also favor progressively closer linkage of the genes in the combination, thus keeping them together as a group over the generations. Another likely difference between the evolution of gene combinations and the evolution of idea combinations is that there may be a greater disparity between the heritability of gene combinations and the transmissibility of idea combinations, since the genes and their combinations are spread by relatively mechanical molecular mechanisms while the ideas of any given set of ideas could become markedly more difficult to spread in combination as the number of elementary ideas being combined increases.
The explosive emergence of new polygenic forms due to the propagation into combination of previously isolated genes might have played a role in causing some of the geologically "sudden" speciation and species changes observed among fossil remains for reasons similar to the mathematical process outlined above for the apparently "sudden" emergence of new idea combinations and their consequences in cultural evolution. This could be regarded a form of genetic innovation arising from continuous quantitative processes bringing genes into new combinations so quickly that the resulting phenotypes appear discontinuous in fossil records. While Eldridge and Gould [1972] use the term punctuated equilibrium to describe the apparently sudden emergences of biological forms, a more fitting term for the process resulting from propagation into combination would be self-punctuating disequilibrium -- as long as it is understood that we are not saying that disequilibrium is punctuated with episodes of equilibrium, but rather, that subtle disequilibrium is punctuated with dramatic disequilibrium. Simpler terms include punctuated gradualness or, (with new word construction) punctuated graduality. (The term punctuated gradualism might be confusing or less precise, since it suggests that an idea or theoretical construct is what is punctuated.) Whatever we call it, we should recognize that even while propagation of genetic (or cultural) varieties continues at extremely low and difficult to observe prevalences, it is not a state of equilibrium even if it appears as such in either live or fossil observations. In the case of biological evolution, propagation into combination depends heavily upon sexual reproduction. The fact that sexual reproduction opens up this avenue to speciation and evolutionary adaptation may even be one of the main evolutionary advantages of sexual reproduction.
What is important for the purpose of the present paper is that the same general mathematical principles may explain both seemingly abrupt biological changes and seemingly abrupt bursts of creativity on a given topic in a population. While propagation of separate replicators may typically be modeled as continuous, their recombination events may be more readily viewed as discontinuities. In the cultural evolution case, the formation of entirely new ideas such as idea Z above may also be considered as discontinuities of evolution. Yet both the enormous numbers of homogenic (replication) events and the more modest numbers of novel or nearly novel heterogenic events forming new combinations of replicators can be viewed as equally discrete or discontinuous events even if their event rates differ by many orders of magnitude. At more fundamental levels of analysis, these events may be understood as abstract representations of hugely complex processes that one might model in terms of continuous physical dynamic functions, for instance, so that one need not assume from the outset that it is fundamentally necessary to posit the existence of physical discontinuities.
The explosive propagation of replicators into new combinations and the sequential accumulation of distinct but synergistic replicators into a combination need not be completely dichotomous phenomena. After the first two of an incrementally synergistic replicator sequence combine, the combination then spreads faster than any of the other still separate but potentially synergistic replicators. That faster propagation for the 2-replicator combination makes the 2-replicator typically shortens the time until the 2-repicator combination forms a 3-replicator combination, and so on through the n-replicator combination and up to the m-replicator combination. A fast-propagating combination may thus start to sweep up the new replicators with which it has not yet synergistically combined, and may even start sweeping up separate new replicator combinations with which it has not yet synergistically combined. The process can therefore become rapidly self-accelerating, to a point that it once again may almost explosively produce the m-replicator combination. In the cultural case, the explosive emergence of the m-replicator combination can also lead to explosive creativity of some new idea such as the one given the general label Z above. When the n-replicator combination is still very sparse in the overall population, any new replicators it encounters may still have less time left to becoming majorities of the (n+1)-replicator combination than does the (n+1)-replicator combination have to reaching majority in the overall population. That can increase the tendency of changes happening in an overall population to happen in muliple-replicator spurts, an effect we might call self-accelerating combinatorial cascading of replicators. Periods of isolation for an n-replicator combination may also give synergistic new replicators at low prevalence in its population more time to spread to majority in the (n+1)-replicator population before the multiple-replicator combination resumes propagation in the overall population. This would further tend to increase the tendency of replicators to attain majority of the overall population in spurts. In the cultural case, it can again increase the explosiveness with which idea Z emerges in a large population.
Practical implications may follow from the above model of population creativity for ideas. For example, proposals to make education highly uniform and enforced by nationwide testing may tend to limit creativity by reducing the variability of combinations of important ideas. Creativity in an organization or a society might alternatively be enhanced by encouraging the acquisition of highly unusual combinations of ideas and fields of learning. Cultural, educational, and experiential diversity might turn out to increase population creativity by increasing the occurrence rates for extremely rare combinations of ideas that could lead to the formation of new ideas. In particular, this might result in higher creative output for universities, research institutions, and other organizations that deliberately strive for a culturally diverse mix of people. Yet even a 1000-fold increase for an idea combination that exists at a prevalence of 10-9 only involves one person in a million, representing only a tiny dent in the prevalence for extremely common combinations of ideas that would form the mainstream of a society or a subculture. Factors such as that might even be investigated as sources of different creativity rates in different countries. Such practical implications also warrant separate papers in their own right. The focus here is on the role of quantitative processes in a population affecting population creativity, and thus the evolution of ideas.
Returning to the evolutionary argument for ideas in general, the most vigorous precursor ideas tend to recombine with more varieties of new ideas, some of which form an even more vigorously propagating idea set in combination with the precursor set. Ideological variation thus feeds new operands into the quantitative processes of natural selection while the quantitative processes give many subsequent qualitative variations an appreciable chance to occur. The two kinds of change continuously feed back on each other to form a genuine process of evolution. A repetition process therefore happens at a higher level than that of individual idea replications to bring about the full effect of evolution through ongoing cycles of variation and selection -- a central feature of all evolutionary replicator theories.
The sort of predictions generated by mathematical models and computer simulations are falsifiable. This provides a general method for attempting to falsify specific thought contagion hypotheses. In general, propagation parameters can be measured over the duration of the time interval being modeled, as can the initial host populations of the ideas under study. If the propagation parameters remain in some specified interval, then the final host population plus or minus a calculable error margin should be measured at the end of that interval. A host population measurement outside those error margins would then falsify the model being used for the specific ideas under study. The procedure can be quite elaborate, as it involves conducting surveys and performing difficult computations.
Yet particular aspects of a specific evolutionary epidemiology of ideas hypothesis can be falsified by less elaborate means as well. If a proposed model for the spread of anti-abortionism hypothesizes that the belief "abortion as wrong" leads adherents to raise more children than non-adherents, then the whole model could be falsified by, for instance, showing that non-adherents raise equal or greater numbers of children. If such data were gathered, and proven representative of whole societies over long time spans, then there would be no need to begin the more elaborate procedures of measuring additional parameters and performing computations.
In some cases, this method can be performed using existing data sets. For example, the National Health and Social Life Survey [Laumann et al 1994] contains data about respondents' attitudes toward various sex acts. It also has data on reproductive histories. So, after making suitable functional definitions based on the questionnaire, one can statistically reanalyze the raw data set to see how much of the variance in reproductive history is attributable to variance in a particular attitude response. Applying the method to the older respondents could provide results that tend to either corroborate or falsify a hypothesis that a particular sex taboo, for instance, increases its hosts' average number of offspring. [note12]
The overall theory would conceivably be falsified by refuting its premises or the logic and mathematics based on those premises. The assertion that the evolutionary epidemiology of ideas is a useful theory is also falsifiable, for instance by showing that differences in propagation parameters are never great enough to account for large-scale ideological shifts, or that the only parameters that ever account for large-scale ideological shift are the "classical" receptivity parameters.
At present, there is no brain scanning device that tells unambiguously whether someone holds a particular belief or not. Nor is there a memory probe that recovers all the communications that led someone to adopt a particular idea. This means that measuring host populations and propagation parameters must depend on survey methodology. Questionnaires and interview protocols may thus form functional definitions of the abstractions used in the evolutionary epidemiology of ideas.
The rates of occurrence of specific events must also be measured for use in a quantitative model, in order to determine the propagation parameters. Surveys already exist which ask how many children people have. Parent to child idea transfer rates would have to be measured by new survey designs that interview both parents and offspring. Human subjects would in general have to be asked to remember what roles their friends or parents may have played in imparting specific beliefs. Respondents "crediting" their parents or people the parents designate on the basis of ideological similarity would be counted toward a idea's parental transmission. Respondents "crediting" other people would be counted toward the idea's non-offspring conversion rate. Much work and proper funding are needed for such surveys.
In a few cases, raw data may have been incidentally gathered for entirely different purposes. The Mormons, for instance, have kept unusually extensive long-term records not only of member births and deaths, but also of how many children followed their parent's faith and how many dropped out, as well as which members arrived through non-parental conversions [Ludlow, 1992]. A project to re-tabulate such data by types of events should enable the calculation of parameters for use in the above differential equations or computer simulations. The resulting host population versus time figures can then be compared to the records of total membership also culled from church records.
With non-human animals, behavior and communication can in principle be monitored with tiny, individual electronic recording devices and studied in detail later. The method is ethically encumbered for humans, but some useful propagation data may still emerge in limited consensual monitoring studies. Though it gets past some of the limitations of surveys, it is still an enormous project.
Small recording devices may also be used in animal experiments in the evolutionary epidemiology of ideas. Bonner has documented the imitational spread in British titmice of the learned skill of pecking through foil caps of milk bottles delivered outside of houses [Bonner, 1980]. A more deliberate experiment could place a large number of smaller artificial food containers throughout an extended (perhaps closed) environment and monitor them. They would be designed so as not to resemble any previous food source, but would be easily opened by the species under study. A container that looks like a distinctive pebble but can be pecked open in a particular spot might do. (To study multiple modes of transmission, care may have to be taken to insure that a single bird cannot have too many imitators in a single feeding, or else the idea will spread to saturate the population under study in less than one generation. Limiting the food per container and choosing a solitary-foraging bird may be necessary.) A large display screen with computer-simulated images of birds opening the containers and eating from them could then be used to release a synthetic idea into a small initial host population. Subsequent monitoring of the birds for reproductive rates, death rates, parent to offspring imitation rates, non-offspring imitation rates, and "forgetting" rates could yield useful parameters that differ between host and non-host populations. Observed host and non-host population sizes versus time can then be compared to those predicted using the quantitative theory and measured parameters.
Human society also affords some specialized environments in which to investigate thought contagions by investigating thought contagion relics. Modern libraries, for instance, allow us to track the proliferating citations of scholarly papers indexed in a computer database.
Some look at the lack of a strictly physical definition of a thought contagion's instantiation as reason for dismissing the evolutionary epidemiology of ideas entirely in favor of theories that describe the same phenomena in terms of genes. Genes, after all, have reliable molecular definitions and are detected using impartial technologies such as nucleotide sequencing. One problem with this argument is that even genetic psychology theories still require functional definitions of psychological features as abstract as an idea. Thus, if an evolutionary psychologist hypothesizes that the valuation of female premarital chastity is genetically based, the empirical investigation must include a method of deciding which females value premarital chastity and which females do not. Then some specific genetic factor must be linked to the presence or absence of this trait. It should, moreover, not just be responsible for learning in general, as the same gene would then account for the ability to learn either the valuation of chastity or the valuation of promiscuity. Rather, it should be a genetic factor whose phenotype and evolutionary adaptation can somehow be more specifically identified with chastity preferences than with strong sexual cravings or cerebral functioning in general.
In many such cases, evolutionary epidemiology of ideas hypotheses are on empirically stronger footing than alternative genetic psychology hypotheses. We can, for instance, observe the hypothesized idea replication event of parents telling sexually eager daughters to "say no." Yet no one has identified and observed a gene that makes daughters want to repress the sexual urges brought on presumably by other genes. The evolutionary epidemiology of ideas hypothesis involves parents feeling economically motivated to impart the idea to daughters and having more grandchildren when they do so Lynch [1996]. It is, in essence, as much Darwinian as the genetic hypothesis is. Yet the evolutionary idea replicator hypothesis is more parsimonious: a competing genetic explanation must still account for the observable communication of sexual repression messages from parents to daughters.
When a communication event is not observed, however, it can serve as indirect evidence of genetic influences. For instance, if parents and others are consistently observed not teaching their daughters any preferences for male lower body physique, but daughters develop their preferences anyway, then a genetic hypothesis for such preferences is on stronger footing than an idea replication hypothesis. Specific genetic factors still need to be identified to completely validate this hypothesis, but it at least does not face a stronger and more parsimonious idea replication alternative.
The evolutionary epidemiology of ideas does face methodological challenges in measuring prevalence and propagation parameters that are more complicated than the functional issues in polling. Yet these should not be seized as arguments for the universal superiority of genetic theories.
The evolutionary epidemiology of ideas can thus be expressed as a type of evolutionary cultural replicator theory, provided that the units of replication are carefully defined. Careful distinctions also need to be drawn between different kinds of idea transition events, so as to recognize that there are many kinds of both replication and non-replication events. The net effect of many kinds of replication events can then be modeled mathematically or computationally. Using measured values of propagation parameters, such models can then be empirically tested against observed data.
A theoretical basis also exists for understanding how the differential
propagation of existing ideas affects the formation rates of new ideas
and new idea combinations that may then go on to become mass replicators
in their own right. The evolutionary contagion of ideas thus includes mechanisms
for both the variation and natural selection of ideas.
My gratitude goes to Carl Wegner for his early support of my evolutionary thought contagion theory work.
My thanks also to Ted Cloak for discussion and correspondence that contributed
to the development of this paper.
1. Dawkins [1976] introduces
the word "meme" in connection with evolutionary cultural replicator theory,
but did not give it a specific definition at that time. Dawkins [1982]
then clarifies that "a meme should be regarded as a unit of information
residing in a brain (Cloak's 'i-culture')" [p. 109]. Several years later,
Dawkins [1986] goes back on this clarification
and states that "memes" exist in various media, only some of which are
brains. This is a bit like saying that "genes" can exist in proteins because
proteinacious replicators called prions have been discovered. Dawkins [1986]
did not offer any reason for the change, cite a source that did give a
reason, or even note that there was a change from 1982. Dennett [1989]
then expands the meaning of the word "meme" even further by treating "meme"
theory as merely a perspective as distinct from a scientific hypothesis
or theoretical framework. Dennett sums up what he calls "the meme perspective"
with the slogan: "A scholar is just a library's way of making another library."
While the slogan expresses the inverted and counter-intuitive thinking
that often arises in evolutionary cultural replicator theory, it further
reduced the specificity of the term "meme" to a point where it might not
be suitable for scientific theory and research. A pithy slogan seems to
have played a larger role in that definition change than any theoretical
or empirical developments of science, and Dawkins's 1986 definition change
was not even identified as such, let alone explained. The changing of definitions
by Dawkins and Dennett may have given others the impression that the two
of them were fumbling for a definition and needed help in the form of additional
proposed definitions. Dawkins may also have incidentally disseminated the
idea of changing the word's definition without any theoretical or empirical
justification, let alone a good justification. Meanwhile, such unexplained
vacillations may have given critics the idea that the definition of terms
was being treated as if it did not matter, creating a widespread sense
that something other than science was being done. In any case, such a profusion
of definitions followed the early vacillations that the word "meme" now
seems less specific than the word "culture," and is associated with numerous
incompatible meanings. Although the word was apparently coined by Dawkins
to popularize Cloak's [1973] theoretical paradigm,
that fact seems to have been forgotten as people eventually began devising
theoretical paradigms to go with the word rather than words to go with
their own drastically different theoretical paradigms.
In 1997, The Oxford English Dictionary apparently
took note of both the early profusion of definitions and the recent shifting
of definitions to come up with a very broad definition for meme. By that
usage, some thought contagions (as defined in section 7 of this paper)
are "memes," some thought contagions are not memes; some "memes" are thought
contagions, and some "memes" are not thought contagions. The OED definition
reads as follows: "MEME: Biol. An element of a culture or a system of behavior
that may be considered to be passed from one individual to another by non-genetic
means, especially imitation." The term "an element of a culture or a system
of behavior that may be considered to be passed from one individual to
another by non-genetic means" is less specific than Cloak's broad 1973
concept of an "elementary self-replicating instruction." But then the last
part of the OED definition "especially imitation" narrows the meaning in
a way that excludes or deemphasizes one of the most important aspects of
evolutionary cultural replicator theory: inculcation. Dawkins [1976]
did speak of "imitation, in the broad sense," but that extremely crucial
point has been left out of the OED definition. The OED's "especially imitation"
clause deflects attention from ideas that spread by causing inculcation
behaviors, since learning by inculcation is not ordinarily what comes to
mind with the term "imitation." The "especially imitation" clause thus
embraces a more conventional view of people acquiring ideas rather than
ideas acquiring people. It suggests people seeing other people's actions,
and then imitating. But people usually think of imitating as something
that mature people, at least, can choose to do or not. Moreover, the term
"especially imitation" with no reference to "the broad sense" allows people
to interpret the term "meme" as referring entirely or mainly to behaviors
instead of ideas. The relative importance of imitation and inculcation
for a given idea or behavior are matters to be determined by scientific
investigation rather than by definition. The OED definition adds new difficulties
without clearing up incompatibilities in the way the word "meme" is used.
2. The fact that cultural evolution theory did not depend on metaphor to genes evolutionary cultural replicator theory did not depend strictly on metaphor and analogy to genes was demonstrated by the symbolic analysis given by Cloak [1973] and later works. Yet even before Darwin's time, various ideas of social evolution existed that clearly could not have arisen from metaphor to Darwin's work [Burrow, 1966]. Indeed, biological evolutionists appear to have adopted the concept of "heredity" by way of metaphor to the cultural practice of inheritance.
5. For an introduction to partially ordered sets, set theory, and metric spaces, see Kaplansky [1972].
6. Colony sizes here come from Hostetler [1974], p. 188, Table 1, which presents demographic charts of a colony branching event in 1969.
7. This argument is spelled out more completely in Lynch [1997].
8.The population age-time profiles [
9. The term "longevity" might be replaced by "durability" to better signify the resistance to dropout and mortality.
10. Homogenic multistage events such as the ones
that add up to
11. These arguments about the likelihood of simultaneous independent innovations should not be naively construed to mean that all claims of independent innovation are accurate. We still need to consider such claims case by case, and to recognize the possibility that informal or forgotten communications originating with an earlier innovator play a part in cases that at first glance appear to be independent re-inventions. No suggestion is made that the priority system for giving credit to innovators be changed.
12. I say "tend to either corroborate or
falsify" because there remains the possibility that a taboo that increased
reproduction for thousands of years no longer does so, or that a taboo
that had no effect for thousands of years suddenly made a difference in
the 1994 data.
[1] Bartholomew, D. J. 1982. Stochastic Models for Social Processes. New York: John Wiley & Sons.
[2] Bonner, J. T. 1980. The Evolution of Culture in Animals. Princeton: Princeton University Press
[3] Boyd, R. and Richerson, P. 1985. Culture and the Evolutionary Process. Chicago: University of Chicago Press.
[4] Burrow J. 1966. Evolution and Society London: Cambridge University Press
[5] Cavalli-Sforza, L., and Feldman, M. (1981). Cultural Transmission and Evolution: A Quantitative Approach. Princeton, NJ: Princeton University Press.
[7] Cloak, F. T. 1966a "A Natural Order of Cultural Adoption and Loss in Trinidad." PhD dissertation, University of Wisconsin at Madison, department of Anthropology. Robert J. Miller, thesis supervisor.
[8] Cloak, F. T. 1966b "Cultural Microevolution." Research Previews 13: (2) p. 7-10.
[7] Cloak, F. T. 1973. "Elementary Self-Replicating Instructions and Their Works: Toward a Radical Reconstruction of General Anthropology through a General Theory of Natural Selection." Paper Presented to the Ninth International Congress of Anthropological and Ethnographical Sciences, Chicago. Scanned version online at http://www.thoughtcontagion.com/cloak1973.htm.
[8] Cloak, F. T. 1975. Is a Cultural Ethology Possible? Human Ecology 3 (3): 161-181]
[9] Cloak, F. T. 1986. The Causal Logic of Natural Selection: A General Theory. In Dawkins, R. and Ridley M. (eds), Oxford Surveys in Evolutionary Biology. Volume 2, 132-186.
[9] Darwin, C. 1859. On the Origin of Species by Means of Natural Selection. London: J. Murray.
[10] Dawkins, R. 1976. The Selfish Gene. Oxford: Oxford University Press.
[11] Dawkins, R. 1982. The Extended Phenotype. San Francisco: W. H. Freeman and Company.
[12] Dawkins, R. 1983. "Universal Darwinism." In Bendall, D. (ed.) Evolution from Molecules to Men, p. 403-425. Cambridge: Cambridge University Press.
[13] Dawkins, R. 1986. The Blind Watchmaker. New York: W. W. Norton & Company.
[14] Dawkins, R. 1989. The Selfish Gene. Revised Edition. Oxford: Oxford University Press.
[15] Dennett, D. 1989. "Memes and the Exploitation of the Imagination," republished in the Journal of Aesthetics and Art Criticism 48:2, Spring 1990, p. 127-135.
[10] Eldridge, N., and Gould, S. 1972. "Punctuated Equilibria: An Alternative to Phyletic Gradualism." In Schopf, T. (ed.) Models in Paleobiology. San Francisco: Freeman, Cooper and Company.
[11] Hamblin, R. 1973. A Mathematical Theory of Social Change. New York: John Wiley & Sons.
[13] Hostetler, 1974. Hutterite Society. Baltimore: The Johns Hopkins University Press.
[14] Kaplansky, I. 1972. Set Theory and Metric Spaces. Boston: Allyn and Bacon, Inc.
[15] Langton, C., Burkhart, R., Lee, I., Daniels,
M., Lancaster, A., 1998. The Swarm Simulation System.
http://www.santafe.edu/projects/swarm
[16] Laumann, E. O., Gagnon, J. H., Michael,
R. T. and Michaels, S. 1994. The Social Organization of Sexuality.
Chicago: The
University of Chicago Press
[17] Ludlow, D. H. ed. 1992. Encyclopedia of Mormonism. New York: Macmillan Publishing Company.
[18] Lynch, A. 1991. "Thought Contagion as Abstract Evolution." Journal of Ideas, 2:1 p. 3-10. Also online at http://www.thoughtcontagion.com/Lynch1991.htm.
[19] Lynch, A. 1996. Thought Contagion: How Belief Spreads Through Society. New York: Basic Books.
[20] Lynch, A. 1997. Thought Contagion and Mass Belief. http://www.thoughtcontagion.com/tcamb.htm
[21] Lynch, A. 1998. "Units, Events and Dynamics in Memetic Evolution." Journal of Memetics - Evolutionary Models of Information Transmission, 2. http://www.cpm.mmu.ac.uk/jom-emit/1998/vol2/lynch_a.html.
[22] Lynch, A. 1999. "The Psychological Underpinnings of Bubbles and Crashes." Talk presented at the meeting of The Institute of Psychology and Markets, at The Aspen Institute, Aspen, Colorado, December 9-10.
[23] Lynch, A. 2000. "Thought Contagions in the Stock Market." Journal of Psychology and Financial Markets 1:1 March 24, 2000, p. 10-23.
[24] Lynch, A. 2001a. "Thought contagion in the stock markets: A general framework and focus on the Internet bubble." Derivatives Use, Trading and Regulation 6:4, p. 338-362.
[25] Lynch, A. 2001b. "Evolutionary Contagion in Mental Software." In Sternberg, R. and Kaufman, J. (eds.), The Evolution of Intelligence. Mahwah, NJ: Lawrence Erlbaum Associates.
[26] Murdoch, G. 1956. "How Culture Changes." In Shapiro, H. (Ed.), Man, Culture, and Society, p. 247-260. Oxford: Oxford University Press.
[27] Rogers, E. M. 1983. The Diffusion of Innovations. New York: The Free Press.
[28] Shiller, R. (2000) Irrational Exuberance. Princeton: Princeton University Press.
[29] Stark, R. 1996. The Rise of Christianity. Princeton: Princeton University Press.
[30] Wilson, D. S., and Sober, E. 1994. "Re-introducing Group Selection to the Human Behavior Sciences." Behavioral and Brain Sciences, 17:585-608.
[31] Witten, M. 1991. "Modeling the Distribution of a 'Meme' in a Simple
Age Distribution Population: I. A Kinetics Approach and Some Alternative
Models." Journal of Ideas, 2:1 p. 19-25.